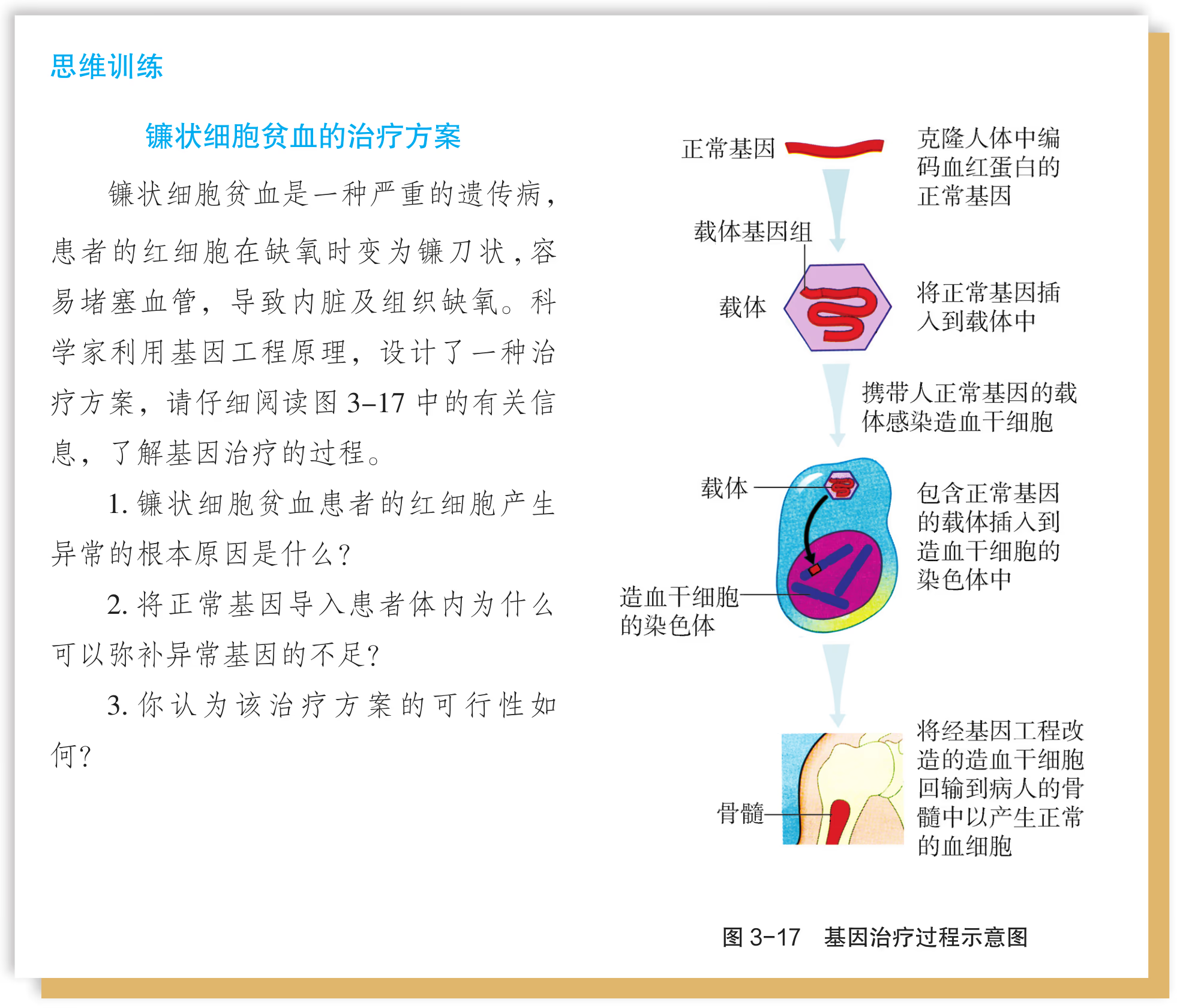
细胞的生命历程
预计阅读时间: 223 分钟细胞分裂
细胞分裂的分类
减数分裂和受精作用与胚胎
减数分裂的过程
胚胎工程的理论基础
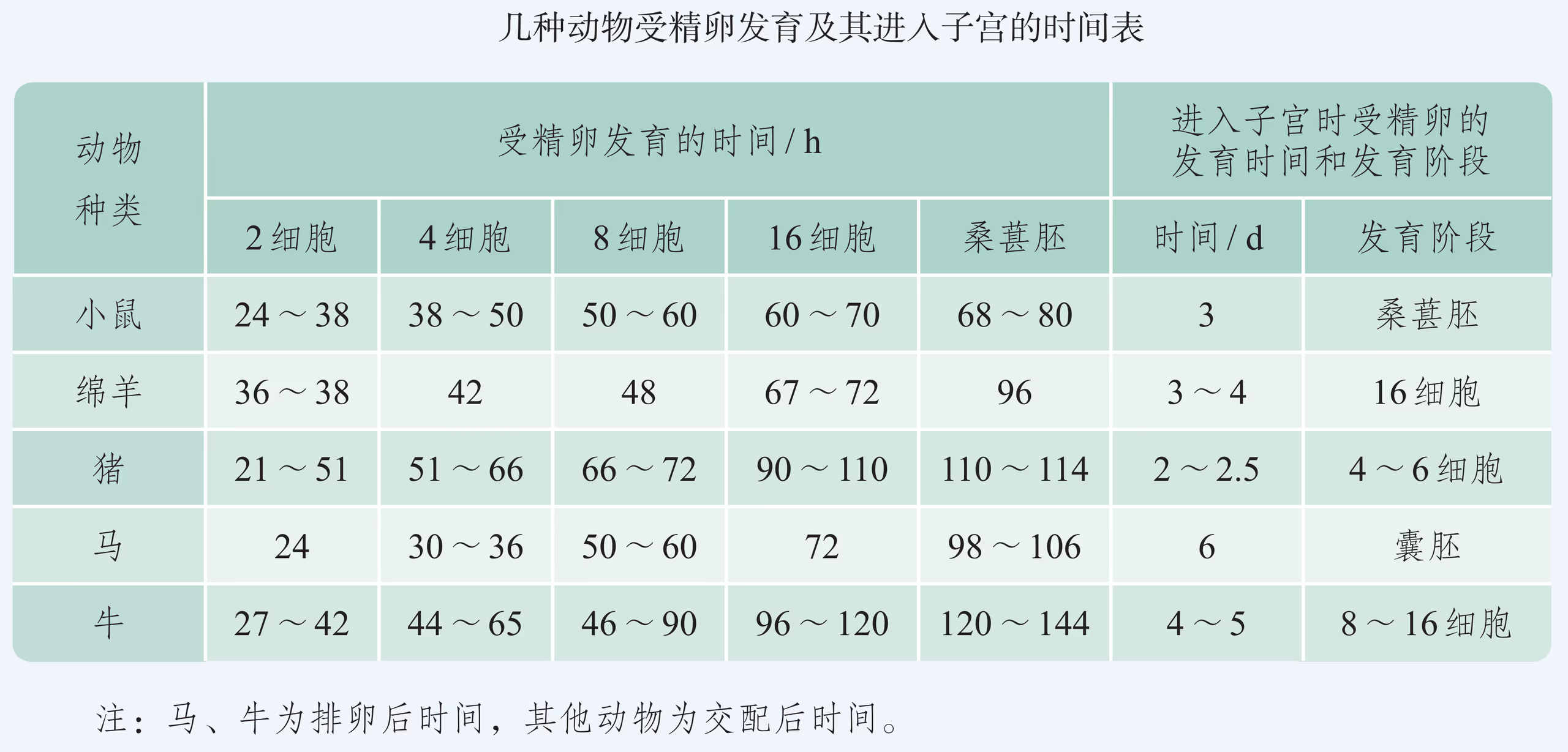
像培育试管婴儿这样,对动物早期胚胎或配子进行显微操作和处理以获得目标个体的技术称为胚胎工程。其操作的主要对象是生殖细胞、受精卵以及早期胚胎细胞。胚胎发育是胚胎工程的理论基础。那么,动物胚胎是如何发育的呢?虽然脊椎动物种类繁多,胚胎发育过程也存在差异,但其早期发育较为相似,分成受精、卵裂、桑椹胚、囊胚、原肠胚与器官形成等阶段(图 2-29)。
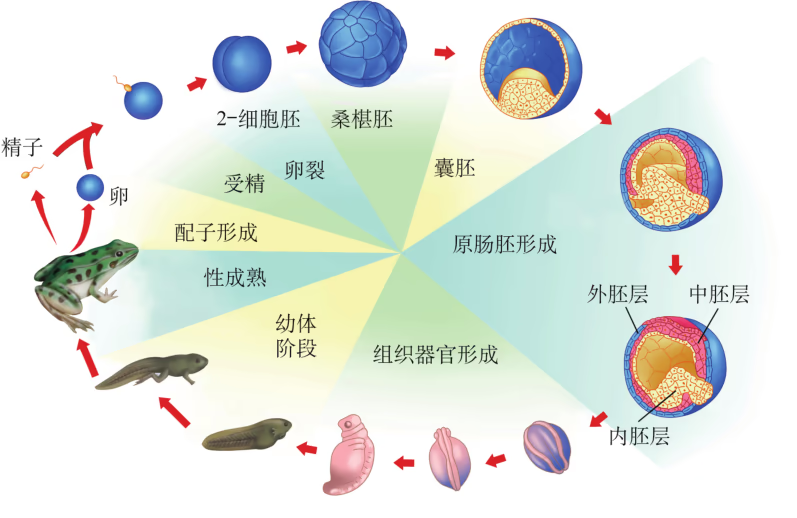
以蛙为例,由于卵黄分布不对称,精子与卵结合之后形成的受精卵分为动物极(发育为外胚层)和植物极(发育为中胚层和内胚层)。受精卵形成后开始卵裂,先分裂成两个细胞,之后细胞通常会逐次倍增,但胚胎的总体积大致不变。受精卵分裂成 个细胞的阶段,称为桑椹胚。在这个阶段,动物极细胞的分裂频率会超越靠近植物极且含有较多卵黄的细胞群,形成中空的球状胚,称为囊胚。随后形成的原肠胚具有外胚层、中胚层与内胚层,这三种胚层在之后会分化出各种组织和器官,逐渐发育成胚胎。
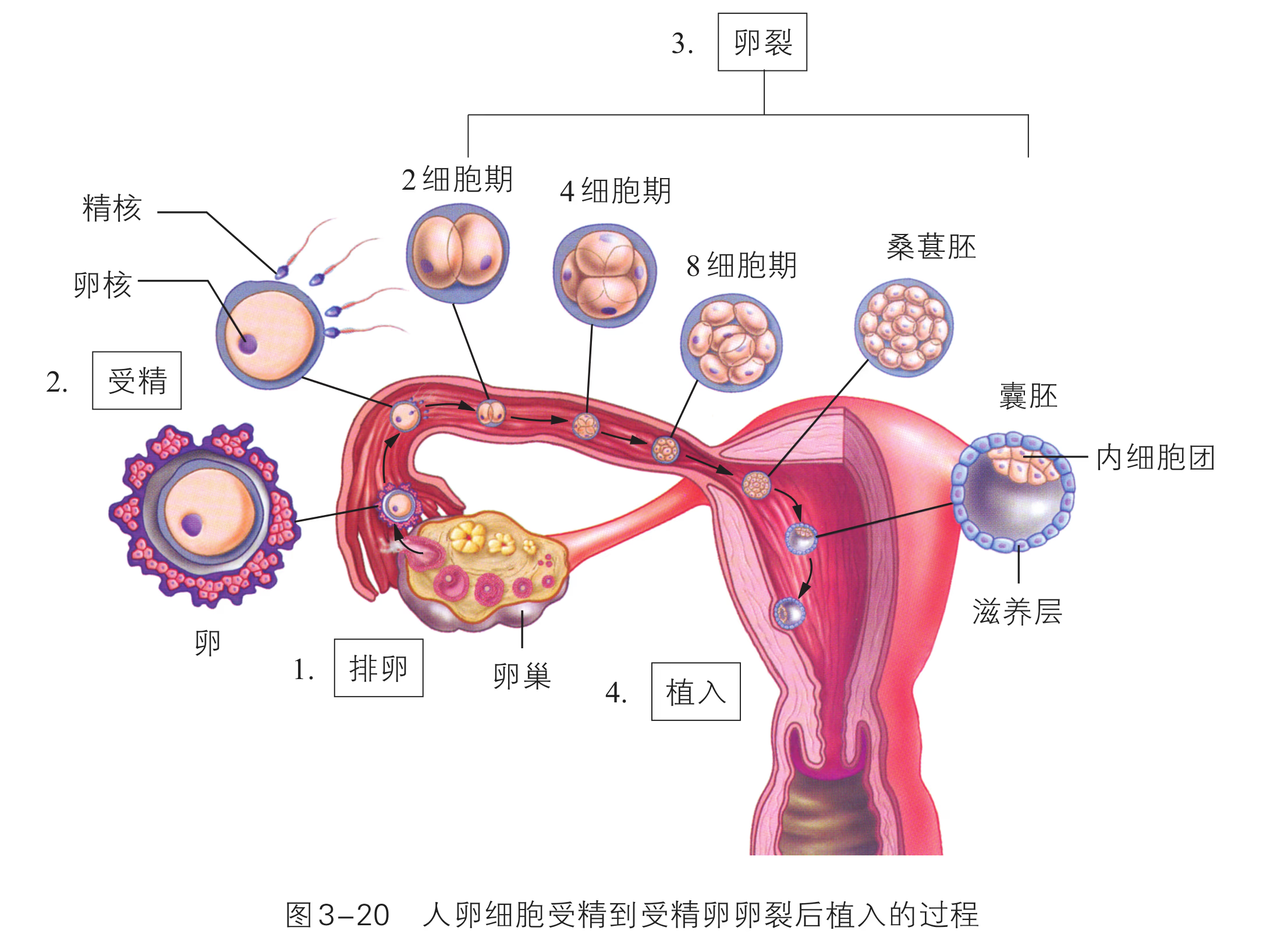
科学家通过模拟自然受孕的过程,逐渐洞悉胚胎发育的每一环节。这些都为胚胎工 程(embryo engineering)的核心技术 --- ---体外受精(in vitro fertilization)和胚胎移植(embryo transfer)等技术的发展奠定了必要的基础,也促进了动物胚胎工程的迅猛发展,拓展了胚胎工 程的应用领域。
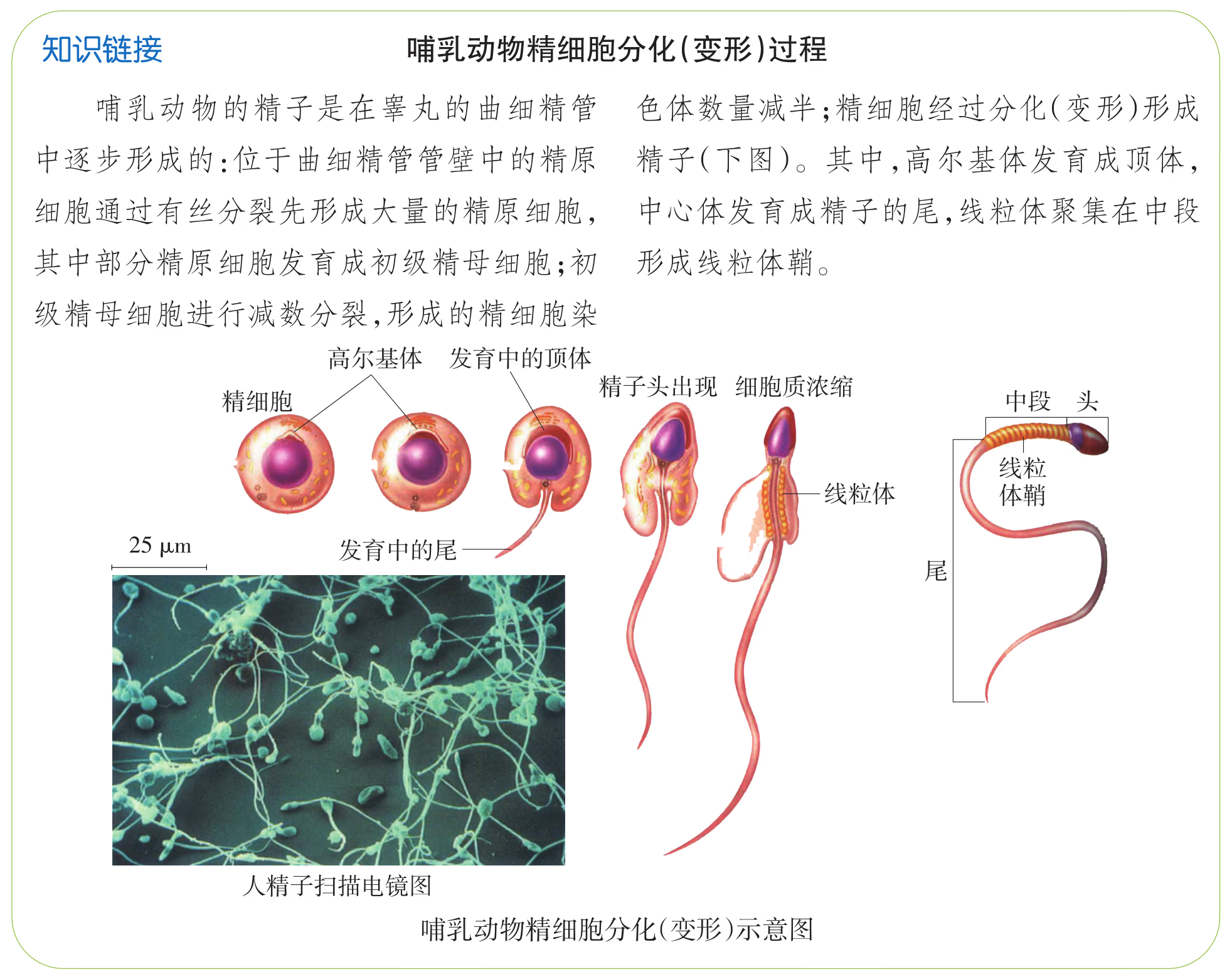
性成熟后,曲细精管内壁上的精原细胞形成初级精母细胞,每个初级精母细胞经过减数分裂形成 4 个精细胞,再经过分化(变形)过程形成精子,每个精子中都含有一个染色体组。精子的发生是依附在支持细胞上进行的,支持细胞为精子的发生提供营养,精子形成后进入曲细精管的管腔中。
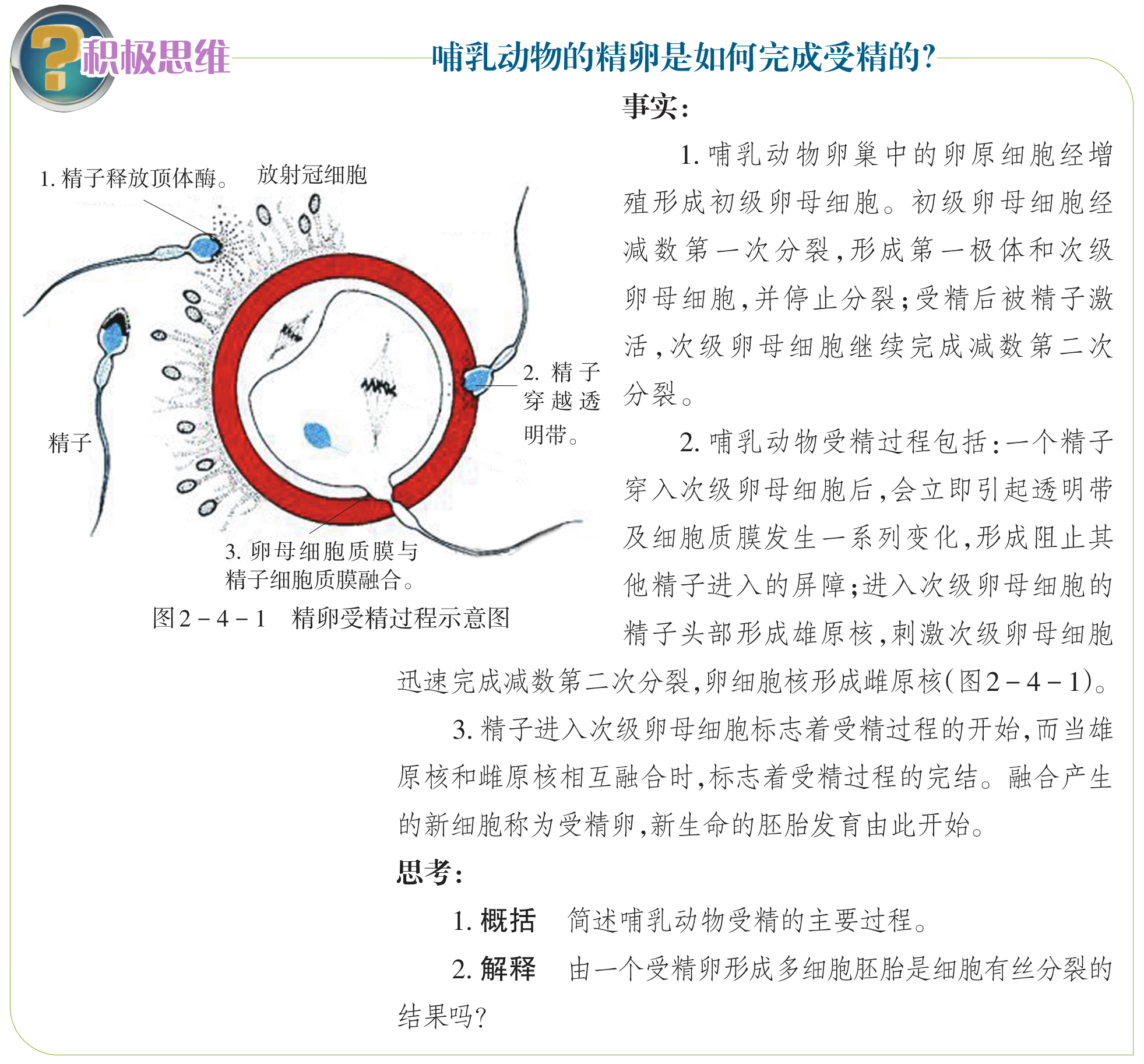
哺乳动物卵细胞的发生是在雌性动物的卵巢中开始的。 一个初级卵母细胞最终可能产生三个极体和一个卵细胞,它们 的细胞中都含有一个染色体组。哺乳动物及其他许多脊椎动 物的卵细胞发育在出生后会停止一段时间,直到雌性动物发 育到性成熟期才继续发育。哺乳动物的初级卵母细胞经过减 数分裂逐步形成成熟的卵子。减数第一次分裂是在排卵前 完成的,排卵后,随着精子进入次级卵母细胞继续减数第二 次分裂,一个次级卵母细胞分裂形成一个成熟的卵子和第二 极体。雌、雄原核逐渐融合,形成受精卵。
当卵细胞质膜和透明带之间出现两个极体、雌雄原核融 合时,表示受精作用已经完成,胚胎发育由此开始。哺乳动物 的胚胎发育是指受精卵发育成幼体的过程,包括桑葚胚 (morula)、囊胚(blastula)、原肠胚(gastrula)等主要发育阶段。
-
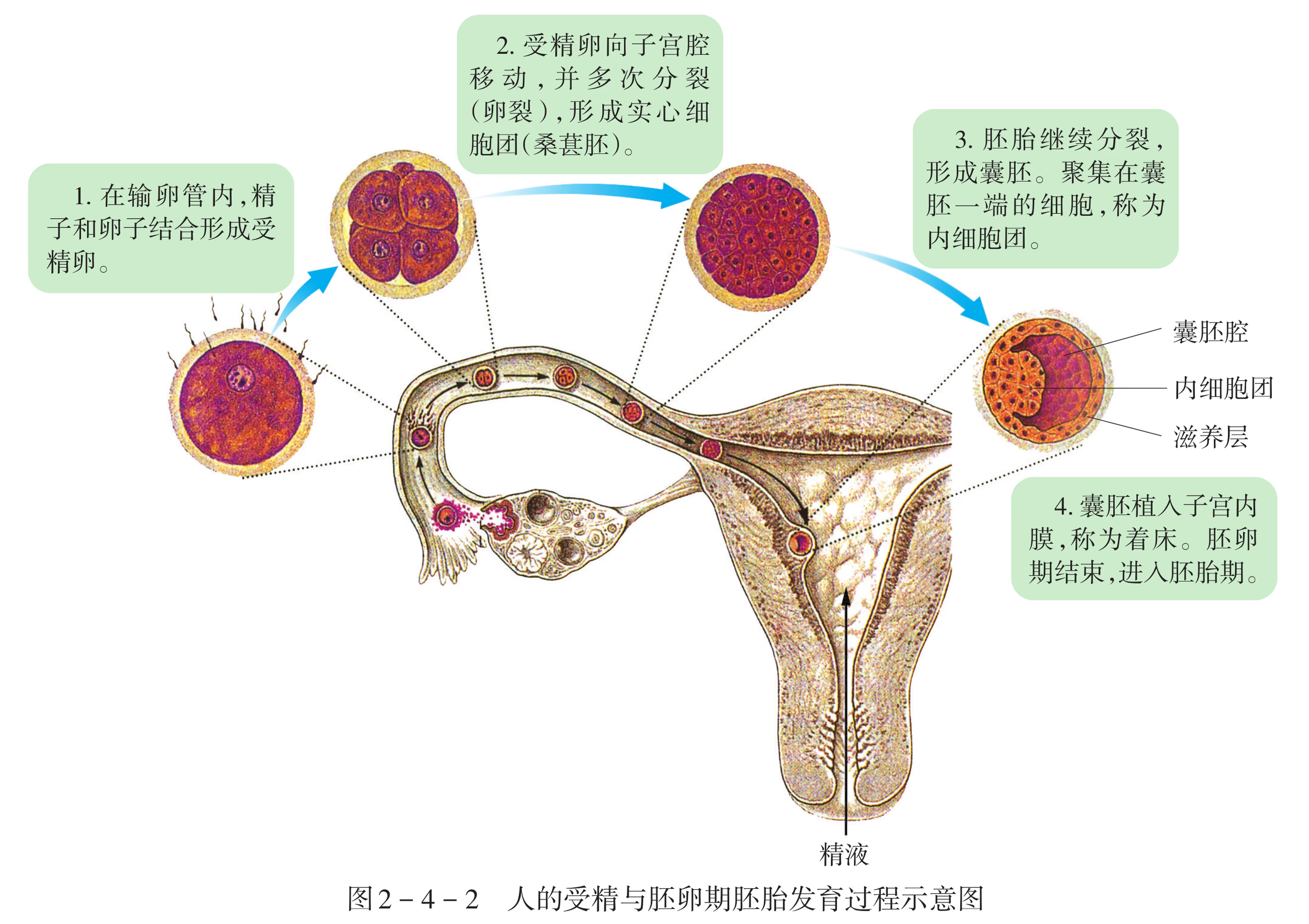
在人的胚胎发育中,受精卵一旦形成,便开始从输卵管移向子宫腔,在移动的同时,受精卵经过细胞分裂,形成囊胚并植入子宫内膜。从受精完成至第 1 周末为胚卵期。在这一时期,胚胎开始着床,囊胚进一步发育。
-
从第 2 周开始至第 8 周为胚胎期。囊胚植入子宫内膜过程中,囊胚壁逐渐发育为滋养层,内细胞团逐渐发育形成二胚层的胚盘,然后继续发育为三胚层的原肠胚。
-
从第 3 周开始,二胚层胚盘中的部分细胞增殖分化形成中胚层。随着三个胚层的分化,三胚层逐渐发育为各种器官的原基。胚盘发育为胚体,胚体凸入羊膜腔,浸泡在羊水 中,胎盘逐渐形成。约在第 3 周末,心脏出现。
-
从第 4 周开始,三胚层逐渐发育,形成眼、耳、鼻等器官的原基;第 4 周末,神经系统开始发生;第 8 周末,各器官原基形成,胚体初具人形,胚胎期发育结束。
-
从第 9 周开始,进入胎儿期,各器官系统继续发育,多数器官逐渐出现不同程度的功能活动;从第 9 周到第 12 周,胎儿的头部仍占优势,躯体长高,脑部继续增长,眼睑闭合,眼内出现视网膜,从外生殖器可辨性别;从第 13 周到第 16 周,胎儿小脑变得突出,一般感觉器官分化,耳郭伸出,出现闭眼和吸吮的动作;从第 17 周到第 20 周,出现睫毛和眉毛,胎毛覆盖皮肤;21 周后,胎儿继续发育直至出生。
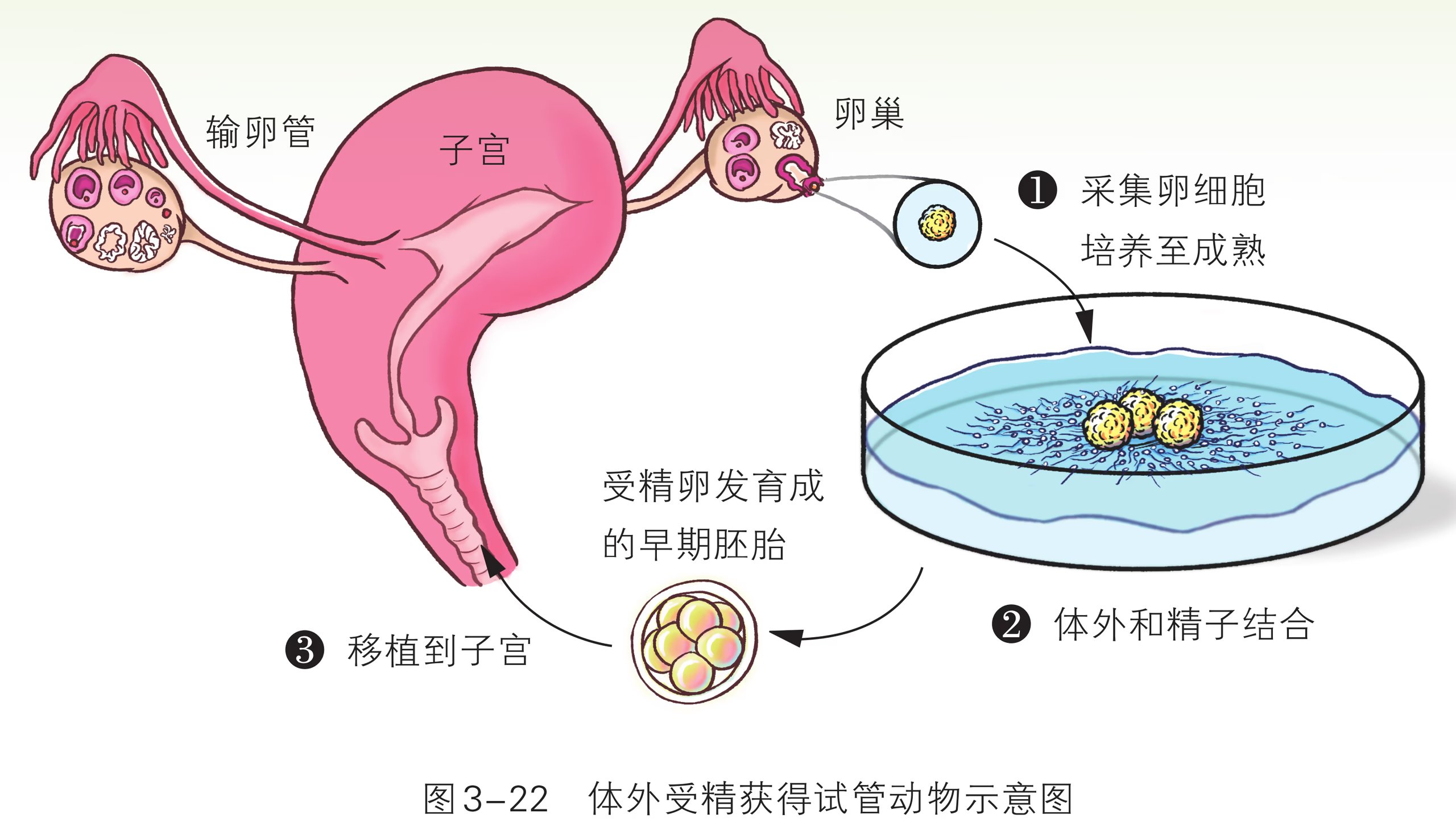
胚胎工程包括体外受精、胚胎移植、胚胎分割移植等技术。体外受精是指哺乳动物的精子和卵在人工控制的环境中完成受精过程的技术。这一技术主要包括卵的采集及成熟培养、精子的采集与体外获能、卵与精子的体外受精等环节。体外受精在动物优良品种的繁育中发挥着极为重要的作用。胚胎移植是将早期胚胎移植到同种且生理状态相同的受体动物体内,使之继续发育成为新个体的技术。胚胎移植实际上是产生胚胎的供体和孕育胚胎的受体分工合作、共同繁育后代的过程(图 2-30)。
胚胎工程的迅猛发展让人们大量繁殖优良家畜品种的愿望,正在成为现实。胚胎工程是指对生殖细胞、受精卵或早期胚胎细胞进行多种显微操作和处理,然后将获得的胚胎移植到雌性动物体内生产后代,以满足人类的各种需求。胚胎工程技术包括体外受精、胚胎移植和胚胎分割等。胚胎工程的许多技术,实际上是在体外条件下,对动物自然受精和早期胚胎发育条件进行的模拟操作。因此,了解哺乳动物受精和早期胚胎发育的规律十分重要。
受精是精子与卵子结合形成合子(即受精卵)的过程,包括受精前的准备阶段和受精阶段。在自然条件下,哺乳动物的受精在输卵管内完成。
-
精子获能:科学研究发现,刚刚排出的精子不能立即与卵子受精,必须在雌性动物的生殖道发生相应的生理变化后,才能获得受精能力,这一生理现象称为"精子获能"。通过对精子获能机制的研究,科学家找到了使精子在体外获能的方法,实现了各种哺乳动物精子在体外条件下的获能,为体外受精技术的建立奠定了重要基础。使精子获能的方法有直接利用雌性动物的生殖道使精子获能;将精子培养在人工配制的获能液中使其获能等。获能液的成分因动物种类不同而有所差异,常见的有效成分有肝素、 载体等。
-
卵子的准备:卵子一般在排出 后才能被精子穿入。动物排出的卵子成熟程度不同,有的可能是初级卵母细胞,如马、犬等;有的可能是次级卵母细胞,如猪、羊等。但它们都要在输卵管内进一步成熟,到 期时,才具备与精子受精的能力。
-
受精阶段:获能后的精子与卵子相遇时,首先它释放出多种酶,以溶解卵细胞膜外的一些结构,同时借助自身的运动接触卵细胞膜。随之,精子的细胞膜与卵细胞膜融合,卵细胞膜外的透明带迅速发生生理反应,阻止后来的精子进入透明带。精子入卵后,卵细胞膜也会立即发生生理反应,拒绝其他精子再进入卵内。精子入卵后,尾部脱离,原有的核膜破裂并形成一个新的核膜,最后形成一个比原来精子的核还大的核,叫作雄原核。与此同时,精子入卵后被激活的卵子完成减数分裂Ⅱ,排出第二极体后,形成雌原核。雄、雌原核充分发育后,相向移动,彼此靠近,核膜消失。这个含有两个染色体组的合子就是受精卵。受精过程结束后,受精卵的发育也就开始了。
多数哺乳动物的第一极体不进行减数分裂Ⅱ,因而不会形成两个第二极体。在实际胚胎工程操作中,常以观察到两个极体或者雌、雄原核作为受精的标志。受精卵形成后即在输卵管内进行有丝分裂,开始发育。胚胎发育早期,有一段时间是在透明带内进行分裂,细胞的数量不断增加,但胚胎的总体积并不增加,这种受精卵的早期分裂称为卵裂。根据胚胎形态的变化,可将早期发育的胚胎分为以下几个阶段。
-
桑葚胚:当卵裂产生的子细胞逐渐形成致密的细胞团,形似桑葚时,这时的胚胎称为桑葚胚。
-
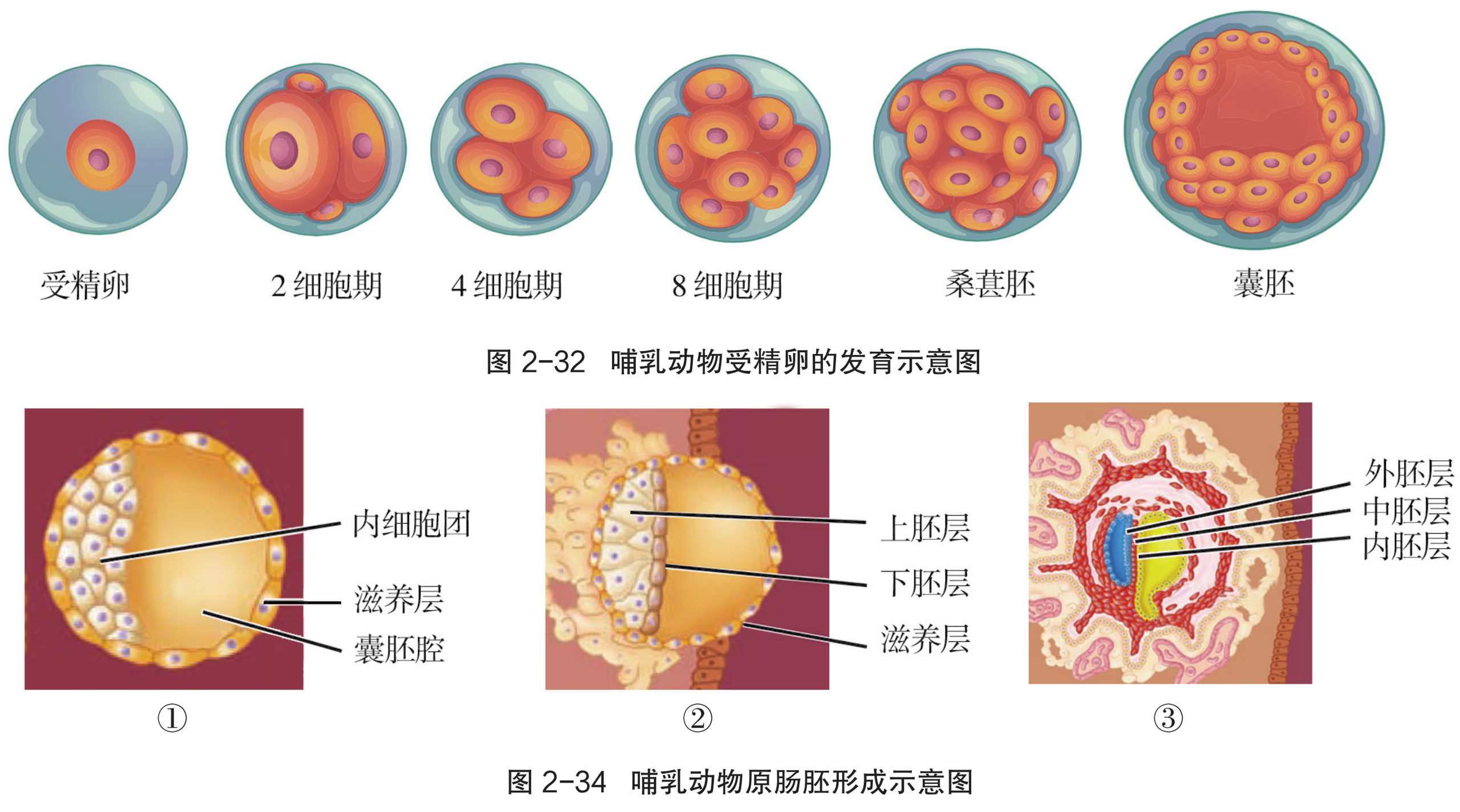
囊胚:胚胎进一步发育,细胞继续分化。聚集在胚胎一端的细胞形成内细胞团,将来发育成胎儿的各种组织;而沿透明带内壁扩展和排列的细胞,称为滋养层细胞,它们将来发育成胎膜和胎盘。
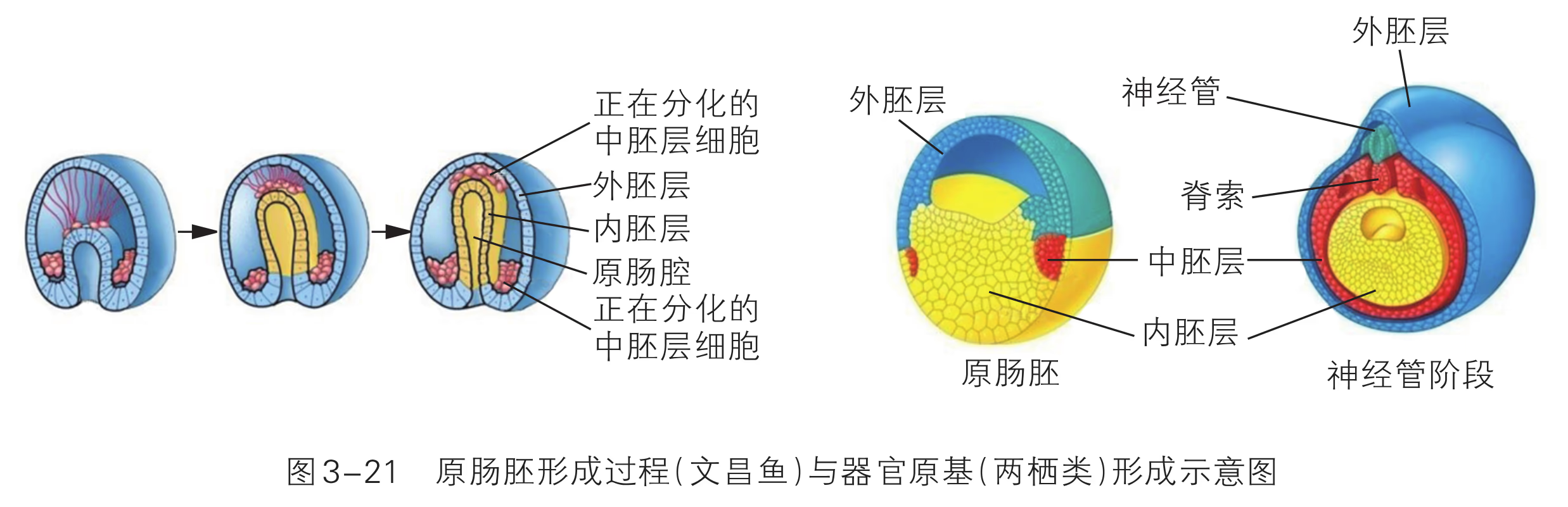
随着胚胎的进一步发育,胚胎的内部出现了含有液体的腔---囊胚腔,这个时期的胚胎叫作囊胚。囊胚进一步扩大,透明带破裂,胚胎从其中伸展出来,这一过程叫作孵化。孵化非常重要,如果不能正常孵化,胚胎就无法继续发育。囊胚孵化后,将发育形成原肠胚。原肠胚表面的细胞层为外胚层,向内迁移的细胞形成内胚层。随着发育的进行,一部分细胞还会在内、外两个胚层之间形成中胚层。这三个胚层将逐渐分化形成各种组织、器官等。
高等动物普遍的生殖方式是有性生殖,雌、雄生殖细胞结合成为受精卵标志着新生命的开始。鼠、牛和羊等哺乳动物的受精方式是体内受精,但新排出的精子不能立即使卵细胞受精,需要在雌性生殖道中发生一系列生理变化后,才具有受精能力,这一生理现象称为精子获能。获能后的精子耗氧量增加,迅速游向卵细胞并与之结合。精子的头部进入卵细胞后不久,精核就与卵核相遇,使彼此的 染色体会合在一起,形成受精卵。
受精完成后,受精卵开始进行快速而有序的有丝分裂,这个过程叫作卵裂(cleavage)。由于卵裂期细胞分裂速度快,胚胎细胞来不及长大,下一次分裂就已开始,所以每个细胞越来越小,但胚胎总体积基本不变。当胚胎细胞达到 16~32 个左右时,形成了致密的细胞团,形似桑葚,叫作桑葚胚。
桑葚胚进一步分裂,胚胎内部形成空隙,逐渐发育成由内细胞团(inner cell mass)、滋养层细胞和囊胚腔构成的囊胚。囊胚中的内细胞团将发育成胎儿的各种组织,而滋养层细胞将发育成胎盘和胎膜。胚胎通过胎盘从母体中获得正常发育所需的氧气和营养物质等。
囊胚进一步发育,内细胞团的细胞不断分裂分化为 2 层细胞。靠近囊胚腔的一层细胞称为下胚层,位于下胚层和滋养层之间的细胞称为上胚层。上胚层的细胞继续分裂分化并迁入下胚层,逐渐置换了下胚层细胞,从而形成一层新细胞,称为内胚层。另一部分上胚层细胞继续分裂扩展,在上胚层和内胚层之间逐渐形成一层新细胞,称为中胚层。形成内胚层和中胚层之后,剩余的上胚层改称外胚层,这样就形成了由 3 个胚层构成的原肠胚。原肠胚的不同胚层之间相互作用,按各自的发育途径分裂和分化为相应的细胞和组织,为形成结构复杂的生命体奠定基础。
哺乳动物从受精卵到幼体的发育过程称为胚胎发育。根据胚胎形态的变化,早期胚胎的发育一般包括三个阶段:桑葚胚、囊胚和原肠胚。这些胚胎细胞还能像受精卵细胞一样发育成完整的个体吗?大量实验表明,在胚胎形成过程中,细胞的分化程度越高,全能性就越弱。尽管桑葚胚和囊胚的细胞在体积和数目上发生了很大变化,但是细胞的功能和发育潜能基本没有改变,分离开来的细胞团仍然可以独立发育成个体。而原肠胚的细胞由于分化程度较高,分割开的胚胎已经不能各自发育成一个新个体。胚胎形成过程中的这些特征为科学家对早期胚胎进行干预和改造提供了可能性。
胚胎分割技术
1942 年,科学家成功地对大鼠二细胞胚胎进行了分离和培养。后来,一些科学家尝试着把绵羊八细胞胚胎分割成 4 份,每份 2 个细胞,再将它们植入母羊子宫内,结果母羊成功地产下 4 只小羊。像这样,用机械方法将早期胚胎平均分割成二分胚、四分胚甚至八分胚,经培养后植入受体,以得到同卵双生或同卵多生后代的技术称为胚胎分割(embryo bisection)技术。这种技术被视为一种胚胎克隆方法。 胚胎分割常需要利用显微操作仪进行操作。在操作时,应选择形态正常、发育良好的桑葚胚或囊胚,在盛有操作液(含有质量分数为 的血清)的培养皿中用分割针或分割刀片将胚胎切开。如果切割的是囊胚,一定要均等分割内细胞团;然后再将吸出的半个胚胎注入一个已经清空的透明带中或直接将其移植到受体中。
- 显微针分割法 该法适用于卵裂球阶段的胚胎。操作时在显微操作仪下将胚胎固定,用显微针在透明带上做一切口,将卵裂球从透明带中移出,并吹吸卵裂球使之离散,将其装入 2 个预先准备好的空透明带内,进行移植。该方法具有较高的同卵双生率,但操作程序复杂。
- 显微刀分割法 该法适用于对桑葚胚和早期囊胚进行分割。在显微操作仪下固定胚胎,用显微刀片将胚胎均匀地一分为二,分别装入空透明带中,进行移植。
- 酶软化透明带后显微分割法 用质量分数为 的链霉蛋白酶溶液软化胚胎透明带。若是紧密化胚胎(如桑葚胚),则需在无钙、无镁的平衡盐溶液中处理 ,使胚胎去紧密化。将胚胎固定,把针置于欲分割的胚胎上面,使其与胚胎呈 ,对准胚胎的正中平分线徐余下压,即可将一个胚胎分割成两半。
- 酶消去透明带后显微分割法 用质量分数为 的链霉蛋白酶溶液消去胚胎透明带,然后用显微针或显微刀将裸胚一分为二。
胚胎分割技术不仅能使可供胚胎移植的胚胎数目成倍增加,而且可以产生遗传性能相同的后代。这对畜牧业生产和科学研究都有重要价值。例如,正常情况下,一头良种奶牛一生约可产犊 10 头,如用胚胎分割技术进行胚胎分割并移植到其他用作"寄母"的普通母牛子宫内,就可以获得大量的良种奶牛。在实际应用中,胚胎分割以二分胚的移植成功率最高。我国相继对猪、牛、马、绵羊、山羊、家兔和小鼠等动物进行了二分胚分割(图 2-4-6),并应用到胚胎移植中。
在动物研究的基础上,英国学者爱德华兹(R. Edwards)和斯特普托(C. Steptoe)最早开展了人类体外受精和胚胎移植技术的研究。1977 年,他们利用腹腔镜取卵,完成体外受精,并在受精卵发育到 8 细胞胚胎阶段移植入母体子宫内获得妊娠。1978 年 7 月,在英国剑桥大学诞生了世界首例"试管婴儿"。我国首例试管婴儿于 1985 年 4 月诞生于台湾。此后,人类的体外受精和胚胎移植技术得到了进一步的发展和应用,为部分生育困难患者带来了福音。
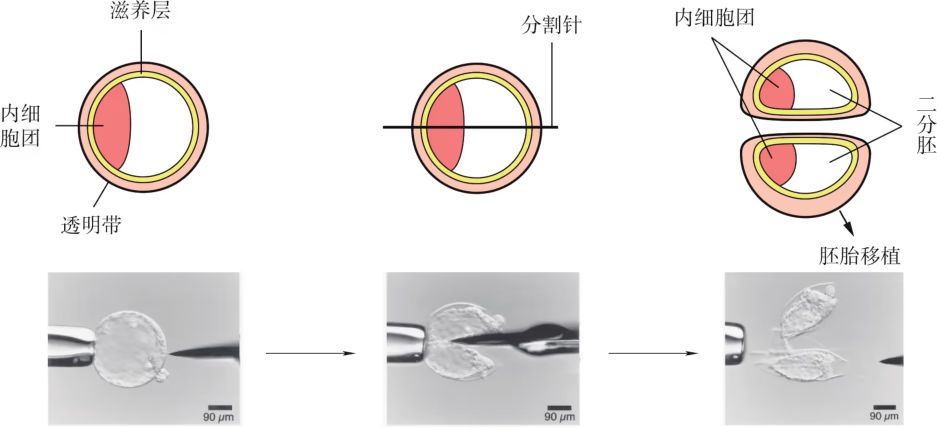
胚胎分割移植是将一枚胚胎用显微手术的方法分割成几份,经体内或体外培养,然后移植入受体中,以得到同卵双生或同卵多生后代的技术。进行胚胎分割时,应选择发育良好、形态正常的桑椹胚或囊胚,将其移入盛有操作液的培养皿中,在显微镜下用分割针或分割刀进行分割。在对囊胚阶段的胚胎进行分割时,还要特别注意将内细胞团均等分割(图 2-31),否则会影响分割后胚胎的恢复和进一步发育。
科学家们首先进行了大鼠卵裂球的分离和培养,随后又以小鼠为实验对象,获得了世界首例半胚来源的活体动物。经过多年的发展,胚胎分割移植在理论和技术方法上都已较为完善,尤其是对牛、羊等大型经济动物实施胚胎分割移植技术,可增加胚胎移植中的有效胚胎数、提高妊娠率,从而增加后代数量。与细胞核移植技术相比,胚胎分割移植具有更高的成功率。
胚胎分割:早期胚胎细胞具有很强的分裂能力,并保持着细胞全能性,能不能将一个胚胎分割成几份,从而提高胚胎的利用率?基于这样的设想,胚胎分割技术逐渐发展成熟。胚胎分割是指采用机械方法将早期胚胎切割成 2 等份、4 等份或 8 等份等,经移植获得同卵双胎或多胎的技术。来自同一胚胎的后代具有相同的遗传物质,因此胚胎分割可以看作动物无性繁殖或克隆的方法之一。胚胎分割所需要的主要仪器设备为体视显微镜和显微操作仪。在进行胚胎分割时,应选择发育良好、形态正常的桑葚胚或囊胚,将它移入盛有操作液的培养皿中,然后在显微镜下用分割针或分割刀分割。在分割囊胚阶段的胚胎时,要注意将内细胞团均等分割。对于不同发育阶段的胚胎,分割的具体操作不完全相同。分割后的胚胎可以直接移植给受体,或经体外培养后,再移植给受体。经过 40 多年的发展,胚胎分割技术日趋成熟。这项技术可以促进优良动物品种的繁殖,产生的遗传性状相同的后代是进行遗传学研究的宝贵材料。此外,在胚胎移植前,进行性别鉴定、遗传病筛查等,对于人工控制动物性别、动物繁育健康后代具有重要意义。
1978 年,科学家将小鼠的桑葚胚一分为二,获得了成功。1979 年,科学家分割绵羊胚胎获得了同卵羔羊。20 世纪 80 年代后,人们建立了系统的胚胎分割方法,并相继得到 ¼ 和 ⅛ 分割胚胎的后代。我国的胚胎工程专家也进行了分割胚胎移植的实验研究,成功地对小鼠、家兔、绵羊、山羊和牛等动物进行了分割胚胎移植,并将二分胚胎分割技术应用到牛和羊的胚胎移植中。尽管胚胎分割技术已经在多种动物中取得成功,但仍存在一些问题,如刚出生的动物体重偏低,毛色和斑纹可能存在差异等。实践证明,采用胚胎分割技术产生同卵多胚的可能性是有限的,分割次数越多,分割后胚胎成活的概率越小。目前,仍然以二分胚胎的分割和移植效率最高,这成为提高家畜胚胎利用率的手段之一。
胚胎移植技术
胚胎工程是指对动物的胚胎进行人为操作和处理以获得目标个体的技术,包括体外受精、胚胎移植和胚胎分割等现代生物技术。 精子和卵母细胞结合后,将受精卵移入相应的培养液中培养。培养液的基本成分包括维生素、氨基酸、蛋白质、无机盐和糖类等。待培养的胚胎发育到特定阶段后,再进行移植。
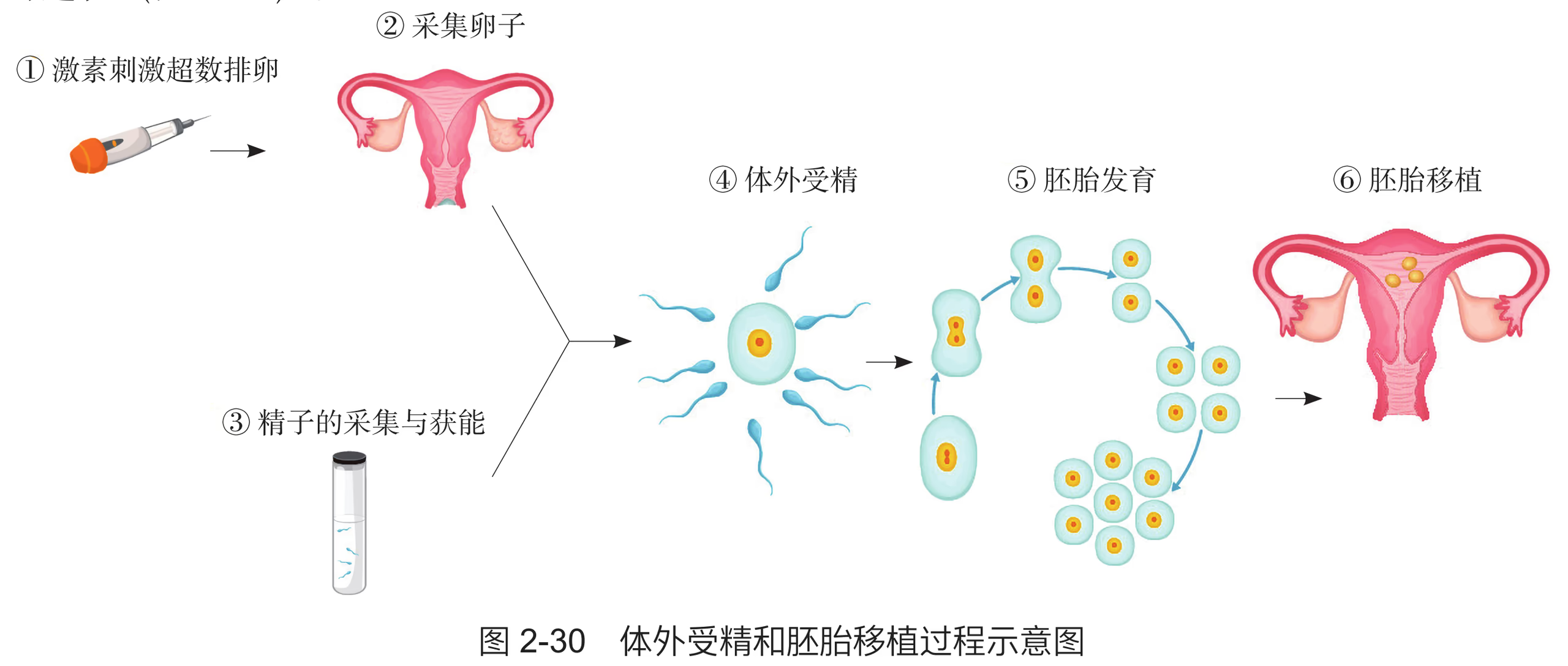
像这样将良种雌性动物配种后,从其生殖道取出早期胚胎或通过体外受精及其他方式(如转基因、核移植)得到的受精卵或胚胎,再移植到同种的同时发情排卵但未经配种的其他雌性动物的体内,使它们进一步发育为新个体的技术,称为胚胎移植技术。 19 世纪末,科学家当时以兔为实验对象成功地完成了哺乳动物的胚胎移植。20 世纪 30 70 年代,胚胎移植成为当时生物学研究的主要方向之一,在多种家畜如绵羊、山羊、猪、牛和马上相继获得成功。20 世纪 60 年代后,科学家开始采用激素对供体进行超数排卵处理,并对受体进行同期发情处理。此后,胚胎移植技术发展迅猛,成为畜牧业中广泛使用的一种技术。 胚胎移植实际上是产生胚胎的供体和孕育胚胎的受体共同繁殖后代的过程。这一技术又被形象地称为"借腹怀胎"。动物胚胎移植过程一般分为供体和受体的选择、供体和受体同期发情处理、供体的超数排卵处理和受精、胚胎收集以及胚胎移植(图 2-4-5)。
胚胎移植作为胚胎工程中的一项技术具有重要的应用价 值。例如,胚胎移植能发挥优良母畜的繁殖能力,迅速扩大优 良畜种数量,加速优良畜种的推广应用;能缩短世代间隔,提 高家畜改良速度;能长期保存冷冻胚胎,便于优良品种的种质 运输和保存。同样,胚胎移植也使胚胎分割和核移植等技术 成为基础生物学研究的重要手段。
胚胎移植是指将通过体外受精及其他方式得到的胚胎,移植到同种的、生理状态相同的雌性动物体内,使之继续发育为新个体的技术。其中提供胚胎的个体称为"供体",接受胚胎的个体叫"受体"。通过任何一项技术(如转基因、核移植和体外受精等)获得的胚胎,都必须移植给受体才能获得后代。以牛的胚胎移植为例,胚胎移植主要包括供体、受体的选择和处理,配种或人工授精,胚胎的收集、检查、培养或保存,胚胎的移植,以及移植后的检查等步骤。
进行胚胎移植的优势是可以充分发挥雌性优良个体的繁殖潜力。在这项技术中,供体的主要职能变为产生具有优良遗传特性的胚胎,繁重而漫长的妊娠和育仔任务由受体取代,这就大大缩短了供体本身的繁殖周期。同时,在对供体施行超数排卵处理后,可获得多枚胚胎,经移植可得到多个后代,这使供体的后代数是自然繁殖的十几倍到几十倍。超数排卵是指应用外源促性腺激素,诱发卵巢排出比自然情况下更多的成熟卵子。
体外受精技术
通过人工操作使卵子在体外受精,经培养发育为早期胚胎后,再进行移植产生的个体。体外受精技术是提高动物繁殖能力的有效措施,还可以为胚胎移植提供可用的胚胎。在这个过程中,首先要做的就是体外受精。哺乳动物的体外受精技术主要包括卵母细胞的采集、精子的获取和受精等步骤。采集到的卵母细胞和精子,要分别在体外进行成熟培养和获能处理,然后才能用于体外受精。一般情况下,可以将获能的精子和培养成熟的卵子置于适当的培养液中共同培养一段时间,来促使它们完成受精。
采集一定发育阶段的精子和卵母细胞是实施体外受精技术的重要环节。 体外受精技术是指哺乳动物的精子和卵子在体外人工控制的环境中完成受精过程的技术。其基本原理是人工模拟体内环境,包括营养、温度、 等,促使卵母细胞成熟,同时使精子获能,最终完成受精作用等。这项技术现已日趋成熟,成为一项重要而常规的动物繁殖技术。体外受精技术包括采集精子、采集卵母细胞和体外受精等主要步骤。
采集精子 采集动物精子的方法主要是假阴道法。假阴道通过模仿发情的雌性动物在交配时阴道的压力、温度和润滑度等条件,创造了适合采精的环境。采精者可手持假阴道,利用假台畜让被采精动物在模拟交配时将精液射入假阴道内,从而达到采集精子的目的。 采集的精子可以立即使用或冷冻保存待用。由于精液中含有精子获能的抑制因子,冷冻液中还含有防冻剂等成分,精液中含有一定量的死精子,一般需要分离并获取有活力的精子。
采集卵母细胞 采集卵母细胞的方法主要有抽吸法、机械破碎法等。 采集较大型的动物如牛、猪的卵母细胞时,一般采用抽吸法。这是一种从活体卵巢中采集卵母细胞时的常用方法。技术人员操作时借助超声波探测仪和内窥镜等进行,将注射器针头刺入卵泡中一并吸出卵泡液和卵母细胞,再在显微镜下进行筛选。该方法的优点是可以有目的地选择卵泡,分离得到适当成熟程度的卵母细胞。活体采卵对于充分发挥优良母畜的繁殖潜力具有一定意义。 采集较小型的动物如小鼠、兔的卵母细胞时,一般采用机械破碎法。这是一种从宰杀后的雌性动物卵巢上采集卵母细胞的方法。技术人员操作时,将刚宰杀的雌性动物体内摘出的卵巢置于培养皿中,用针头刺破卵巢和卵泡,再在显微镜下对卵母细胞进行筛选。该方法的优点是可以得到各种成熟程度的卵母细胞,缺点是在刺破卵巢和卵泡的过程中有可能刺伤卵母细胞。对于较小型的动物,还可以采用促性腺激素处理的方法,使其超数排卵,这样获得的卵母细胞可以直接与获能后的精子进行体外受精。 如果采集到的卵母细胞尚未成熟,需通过体外培养使之成熟。经过体外培养后,一般要通过相关的形态观察指标(如 后的囊胚发育率)对卵母细胞的体外成熟情况进行评价。
人卵泡的发育 人卵泡的形成和在卵巢中的储备是在胎儿期完成的。人的胚胎性别分化后,雌性胎儿卵巢内的卵原细胞经过有丝分裂增加其数量,其中部分卵原细胞发育为初级卵母细胞。由于它们被卵泡细胞所包围,一般称之为卵泡。 新生儿(女性)出生时卵巢内约有 万 万个卵泡。青春期以后,卵泡逐渐减少,女性一般只有 个卵泡能发育成熟,其余卵泡均发育到一定程度即自行退化。 有些哺乳动物和人一样,卵泡的发育也始于胚胎时期。一般认为,哺乳动物出生后卵巢内卵泡不可更新,但仍有科学家在不断研究卵巢中是否存在生殖干细胞。这可能对生殖医学以及干细胞研究等具有深远意义。
体外受精 哺乳动物的交配往往发生于发情的开始或发情盛期,排卵则发生于发情结束前后,这既保证了精子能先于卵母细胞到达受精部位,也给精子留了充足的获能时间。精子获能后发生膜流动性的增加、膜对 通透性提高、胞内环磷酸腺苷(cAMP)浓度的升高、表面电荷降低以及游动方式等一系列变化,其重要意义在于增强了精子的代谢能力和运动能力。 获能后的精子会引起 内流到精子内,同时伴随精子细胞质膜通透性的改变和胞内 升高,刺激顶体反应的发生。顶体反应主要发生在精子头部。精子与卵子利用各自表面的特异性蛋白进行识别后,发生特异性结合。精子头部前端与卵母细胞透明带接触后,顶体外膜与精子头部的细胞质膜融合,顶体破裂释放出的顶体酶溶解卵母细胞周围的放射冠和透明带,精子即可穿过透明带到达卵母细胞表面。随着精子进入卵母细胞,雄原核与雌原核融合,最终形成受精卵。
家兔、猪等动物的精子,放入人工配制的获能液中,经培养即可获能;牛、羊等动物的精子,可放入一定浓度的肝素或相应的 载体溶液中,诱导精子获能。 在人为控制的适宜条件下和专用的受精溶液中,获能的精子和培养成熟的卵母细胞放在一起培养一段时间,可以在体外完成受精过程。
日趋成熟的体外受精技术可用于研究哺乳动物生殖细胞的发生、受精和早期胚胎发育机制等。在家畜品种改良探索过程中,体外受精技术为胚胎生产提供了经济而高效的方法,对充分利用优良种质资源、缩短家畜繁殖周期、加快品种改良速度等具有重要价值。
植物细胞工程
什么是细胞工程
生物技术是应用生命科学的研究成果,对生物或生物的成分、产物等进行改造和利用的技术。生物技术是一个综合性的技术体系,我们可以将它与工程学原理相结合,来进行研究、设计和加工生产,为社会提供服务。细胞工程是指应用细胞生物学、分子生物学和发育生物学等多学科的原理和方法,通过细胞器、细胞或组织水平上的操作,有目的地获得特定的细胞、组织、器官、个体或其产品的一门综合性的生物工程。
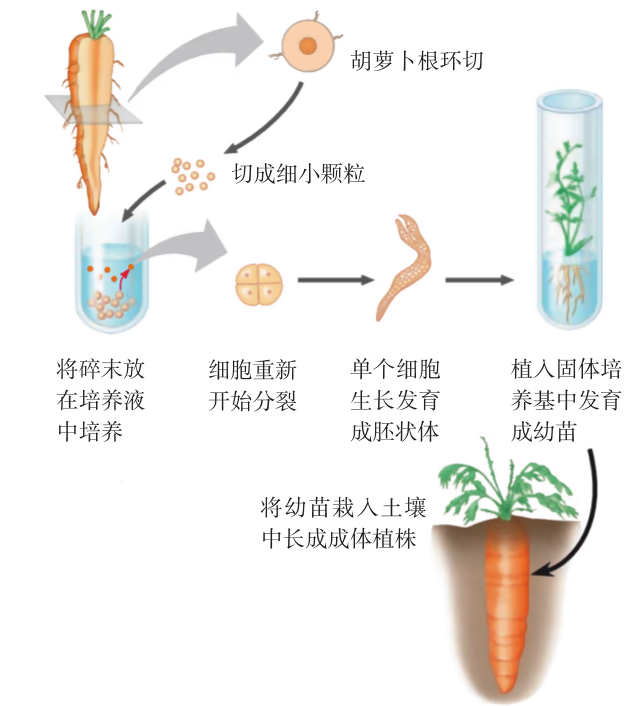
在细胞学说的基础上,德国植物学家哈伯兰特(G. Haberlandt)于 1902 年提出了植物细胞具有全能性的假设,即离体的植物细胞在合适的条件下能够发育成为完整的植株。遗憾的是,受到当时技术和设备条件的限制,哈伯兰特没能证实自己的假设。1958 年,美国科学家斯图尔德(F. C. Steward)经过十几年的艰苦探索,用野生胡萝卜的根韧皮部组织通过组织培养的方法培育出具有根、茎、叶的完整植株(图 2-2)。1965 年,科学家再用分离出的单个胡萝卜细胞进行培养,也实现了整株植株的再生。从哈伯兰特提出植物细胞全能性的假设,到这一假设得到科学实验的证实,经过了半个多世纪。
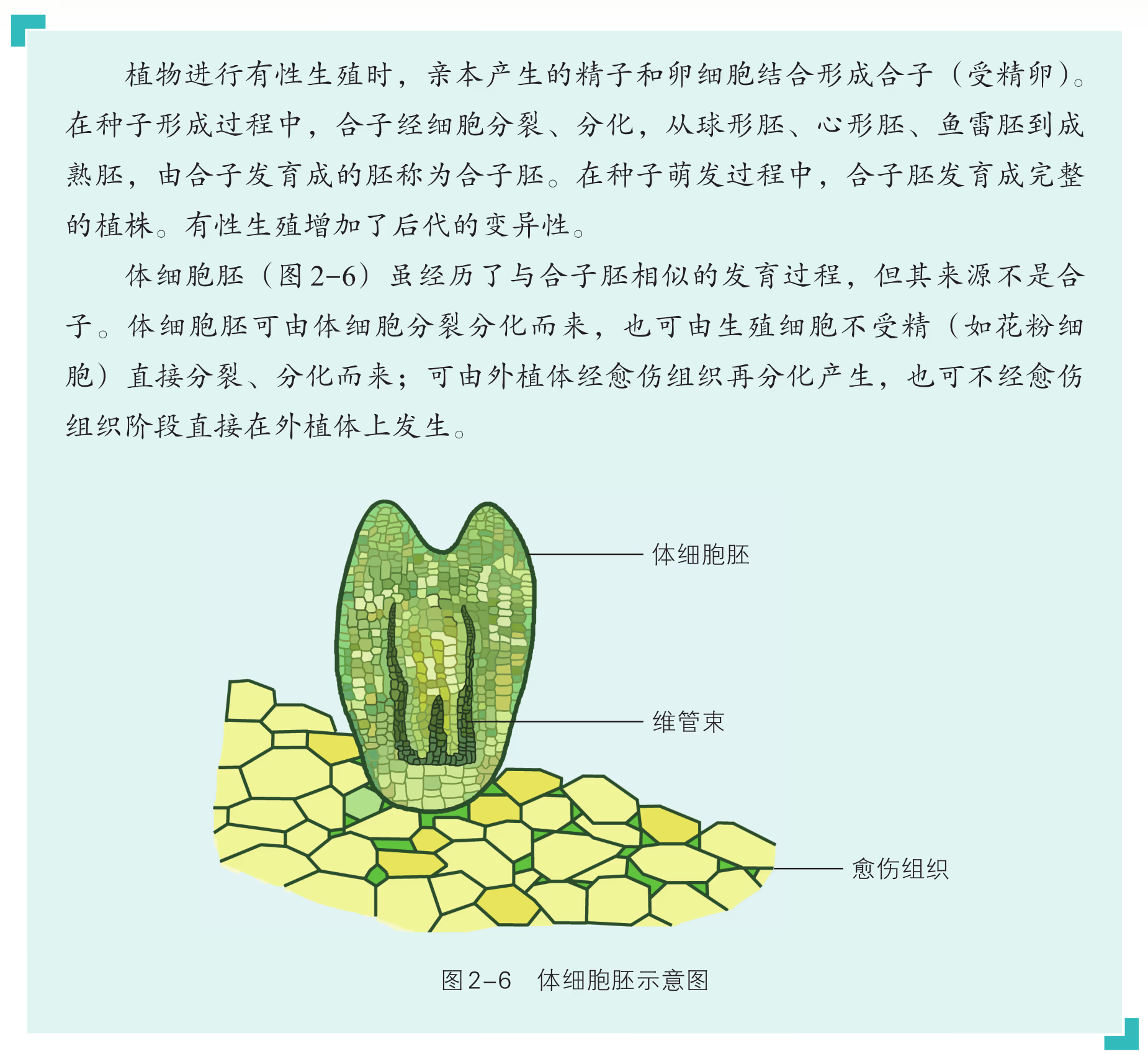
我们已经知道,分化后的细胞虽然在形态、结构与功能上与最初的受精卵存在着显著差异,但仍包含着与受精卵相同的遗传信息。在一定的条件下,分化后的细胞重新获得类似于胚胎细胞一样的分裂和分化能力的过程称为脱分化(dedifferentiation)。 脱分化后的细胞经过细胞分裂,会形成无组织结构的、松散的细胞团,即愈伤组织(图 2-3)。愈伤组织在一定条件下可以重新分化为各种类型的细胞,并进一步发育成完整的植株,这个过程称为再分化(redifferentiation)。
基于植物细胞的全能性理论,将离体的植物细胞、组织或器官(外植体)在适宜的培养条件下经脱分化形成愈伤组织,然后经过再分化,形成芽、根并最终发育成完整植株,这一过程称为植物组织培养。
在植物组织培养中,细胞的脱分化和再分化是两个关键步骤。多种内部因素(如植物遗传性状和生理状况等)和外部因素(如营养和环境条件等)会影响这两个步骤。在这些影响因素中,生长素和细胞分裂素的浓度比例起着关键作用,它们对愈伤组织的生长和分化产生不同的影响(图 2-4)。
植物细胞工程的原理
细胞经分裂和分化后,仍然具有产生完整生物体或分化成其他各种细胞的潜能,即细胞具有全能性。但是,在生物的生长发育过程中,并不是所有的细胞都表现出全能性,比如,芽原基的细胞只能发育为芽,叶原基的细胞只能发育为叶。这是因为在特定的时间和空间条件下,细胞中的基因会选择性地表达。植物组织培养是指将离体的植物器官、组织或细胞等,培养在人工配制的培养基上,给予适宜的培养条件,诱导其形成完整植株的技术。这些离体培养的植物器官、组织或细胞被称为外植体。
-
植物细胞一般具有全能性。在一定的激素和营养等条件的诱导下,已经分化的细胞可以经过脱分化,即失去其特有的结构和功能,转变成未分化的细胞,进而形成不定形的薄壁组织团块,这称为愈伤组织。愈伤组织能重新分化成芽、根等器官,该过程称为再分化。植物激素中生长素和细胞分裂素是启动细胞分裂、脱分化和再分化的关键激素,它们的浓度、比例等都会影响植物细胞的发育方向。将愈伤组织接种到含有特定激素的培养基上,就可以诱导其再分化成胚状体,长出芽和根,进而发育成完整的植株。
-
植物细胞外面有一层细胞壁,这层细胞壁阻碍着细胞间的杂交。因此,在进行体细胞杂交之前,必须先利用纤维素酶和果胶酶去除这层细胞壁,获得原生质体。杂交过程中的一个关键环节,是原生质体间的融合,这必须要借助一定的技术手段才能实现。人工诱导原生质体融合的方法基本可以分为两大类---物理法和化学法。物理法包括电融合法、离心法等;化学法包括聚乙二醇(PEG)融合法、高 ---高 融合法等。融合后得到的杂种细胞再经过诱导可形成愈伤组织,并可进一步发育成完整的杂种植株。
-
植物体细胞杂交是指将不同来源的植物体细胞,在一定条件下融合成杂种细胞,并把杂种细胞培育成新植物体的技术。利用这项技术,科学家培育出了白菜---甘蓝、普通小麦---长穗偃麦草等杂种植株。植物体细胞杂交技术在打破生殖隔离,实现远缘杂交育种,培育植物新品种等方面展示出独特的优势。
植物细胞工程的应用
植物细胞工程在农业、医药工业等方面有着广泛的应用,并且取得了显著的社会效益和经济效益。
-
快速繁殖:将优良植株的茎尖或叶片等器官进行离体培养,在短时间内可以获得大量遗传性状一致的植株,这种技术称为植物快速繁殖技术。继兰花的快速繁殖实现后,该技术得到了快速发展,其适用对象从最初的观赏植物逐渐发展到果树、林木、蔬菜和田间作物。现已有上千种植物可通过组织培养技术进行快速繁殖。我国科学家屠呦呦团队还利用组织培养技术,建立了青蒿国家种质资源库,用于检验不同种质资源的青蒿素含量、耐旱性、抗病虫害等指标,以便寻找出更优质的品种。
-
作物脱毒:利用植物组织培养技术还可以进行植物脱毒(即降低或去除病毒对植株的感染)。目前已知可以感染植物的病毒有数百种,受到感染的植株通常会出现病斑,其正常生长受到严重影响。绝大多数的病毒是通过植物体内的维管组织传播的,而位于种子植物胚珠中央的珠心组织与周围其他组织没有维管联系,因此通过珠心组织培养可以获得无毒植株。此外,通过离体培养分裂旺盛的茎尖也可以脱除植物病毒。
-
种质保存:植物细胞全能性的发现和证实还为植物种质资源的长期保存开辟了一条新途径。许多植物的组织培养物在液氮中超低温保存以后,仍能保持很高的存活率而再生出新植株,并保持原来的遗传特性。例如,胡萝卜和烟草的悬浮细胞超低温保存 6 个月以后仍然能恢复生长并分化出植株。超低温保存的植物材料,可以是植物的悬浮细胞,也可以是愈伤组织或胚状体,还包括原生质体、花粉、幼胚、芽或茎尖分生组织等。超低温保存植物材料的原理与保存动物材料的原理一样,在 -196℃ 的条件下,活细胞内的物质代谢和生命活动几乎完全停止,细胞、组织、器官在超低温保存过程中不会引起遗传性状的改变,也不会丧失形态发生的潜能。在植物自然资源受到过度开发和严重破坏的情况下,利用人工种质库长期保存珍稀的濒危植物更加迫在眉睫。同时,这样的方法比保存植物的种子更为有利和经济,还能在需要的时候迅速且大量地繁殖植物。
-
单倍体育种:常规选育出一个可以稳定遗传的作物优良品种,一般要经过 年的连续筛选。而单倍体育种可以先通过花药(或花粉)培养获得单倍体植株,然后经过诱导染色体加倍,当年就能培育出遗传性状相对稳定的纯合二倍体植株,这极大地缩短了育种的年限,节约了大量的人力和物力。单倍体育种已成为作物育种的一条有效途径。此外,由于大多数单倍体植株的细胞中只含有一套染色体,染色体加倍后得到的植株的隐性性状容易显现,因此它也是进行体细胞诱变育种和研究遗传突变的理想材料。
20 世纪 20 年代,科学家首次在自然界发现高等植物曼陀罗的单倍体植株,它与正常二倍体植株相比,叶小、株矮、生存能力弱且高度不育。但是,单倍体的发现对如何在育种中缩短育种周期、获得纯系植株等具有重要意义。通过常规的杂交育种(cross breeding)方法培育新品种,一般需要花费较长时间才能筛选出具有稳定遗传特性的新品种。而通过单倍体育种(haploid breeding)方法培育新品种,可以大大缩短育种的时间,且可以获得具有稳定遗传特性的优良品种,节约了人力物力。单倍体的诱导与利用是植物细胞工程成功应用的典型范例。单倍体的育种思路是,先通过培育小孢子,获得单倍体植株幼苗,再诱导其染色体数目加倍,从而获得纯合二倍体植株。
成熟的花药由花药壁和花粉囊及其中的花粉等结构组成。经过适当的诱导,花粉囊中的花粉可发育成单倍性的胚状体或愈伤组织,最终形成单倍体植株。在诱导单倍体植株的某些阶段(如形成愈伤组织时期或幼胚时期),一般用质量分数为 的秋水仙素溶液处理 ,然后按常规途径培养,就可能获得染色体数目加倍的,能正常生长发育并开花、结果的二倍体植株。
-
生产次生代谢产物:植物细胞除了能诱导形成完整植株外,还能像微生物一样在生物反应器里进行大规模的悬浮培养,用于生产重要的植物次生代谢产物。次生代谢产物是一类细胞生命活动非必需的小分子有机化合物,其产生和分布通常有种属、器官、组织以及生长发育时期的特异性。
初生代谢是生物生长和生存所必需的代谢活动,因此在整个生命过程中它一直进行着。初生代谢物有糖类、脂质、蛋白质和核酸等。次生代谢不是生物生长所必需的,一般在特定的组织或器官中,并在一定的环境和时间条件下才进行。植物代谢会产生一些一般认为不是植物基本的生命活动所必需的产物---次生代谢物。次生代谢物是一类小分子有机化合物(如酚类、萜类和含氮化合物等),在植物抗病、抗虫等方面发挥作用,也是很多药物、香料和色素等的重要来源。
由于植物细胞的次生代谢物含量很低,从植物组织提取会大量破坏植物资源,有些产物又不能或难以通过化学合成途径得到,因此人们期望利用植物细胞培养来获得目标产物,这个过程就是细胞产物的工厂化生产。植物细胞培养是指在离体条件下对单个植物细胞或细胞团进行培养使其增殖的技术。它不占用耕地,几乎不受季节、天气等的限制,因此对于社会、经济、环境保护具有重要意义。
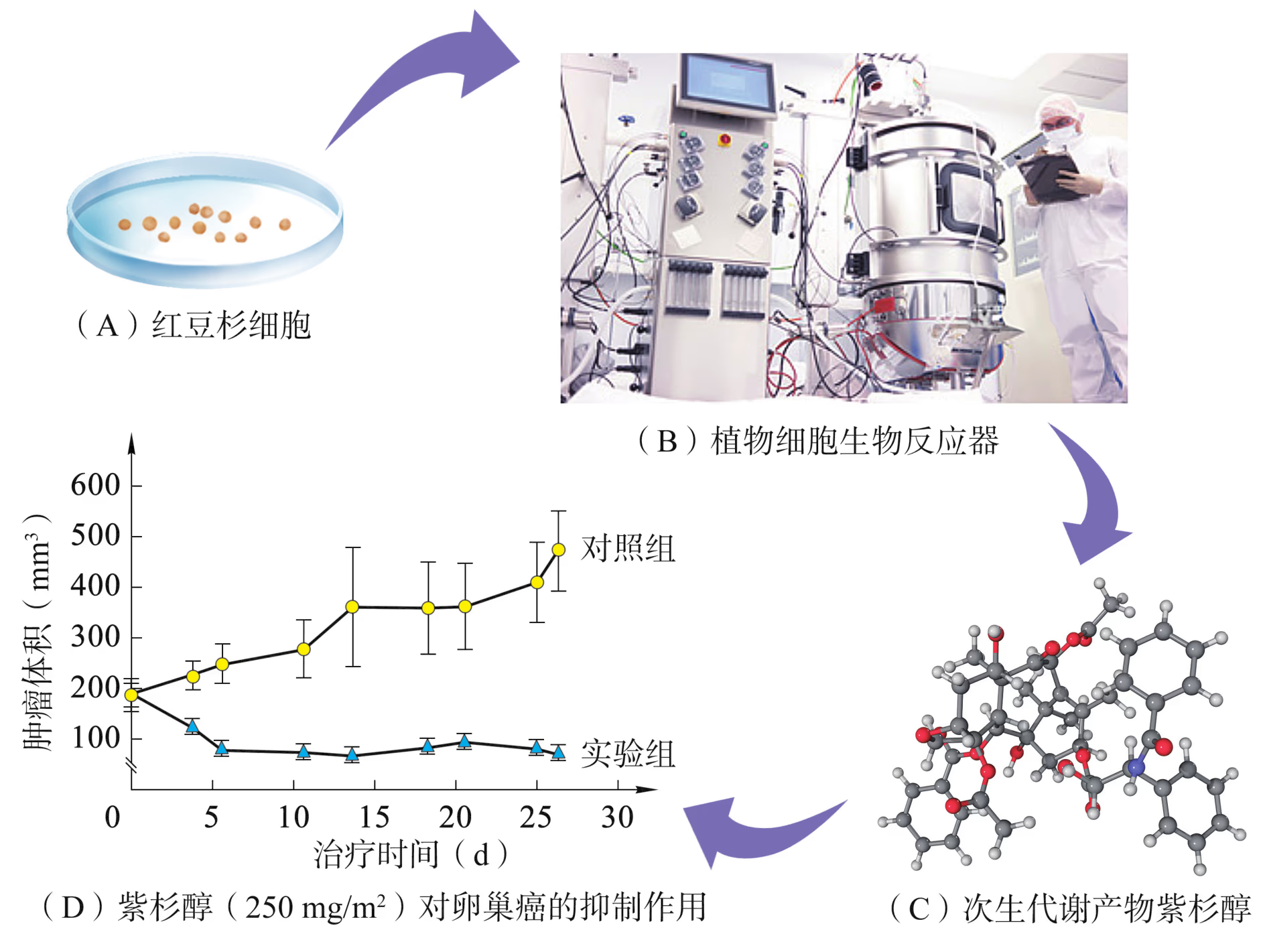
例如,紫杉醇是红豆杉属植物中的一种次生代谢产物,它是迄今为止发现的最好的天然抗癌药物之一。红豆杉属植物树皮中紫杉醇的含量极低,而且树种生长缓慢。通过剥树皮来提取紫杉醇,会导致红豆杉大量死亡(图 2-7)。所以,紫杉醇的产量受到很大限制。为了满足市场对紫杉醇的大量需求,同时减少对自然资源的破坏,科学家们正在尝试各种技术路线。其中,植物细胞培养技术可以解决这一难题。通过对红豆杉细胞的悬浮培养,可以从中分离出高纯度的紫杉醇。
突变体的利用:在植物的组织培养过程中,由于培养细胞一直处于不断增殖的状态,因此它们容易受到培养条件和诱变因素(如射线、化学物质等)的影响而产生突变。从产生突变的个体中可以筛选出对人们有用的突变体,进而培育成新品种。
植物体细胞杂交技术
植物体细胞杂交又称为植物原生质体融合,是指将不同来源的植物细胞在一定条件下融合形成杂合细胞,并使之分化再生,最终形成新植物体的技术。例如,白菜和甘蓝是两种蔬菜,存在种间生殖隔离。育种专家借助植物体细胞杂交技术培育出了白菜---甘蓝这一新品种(图 2-9)。与动物细胞不同的是,植物细胞外周覆盖着一层致密而坚硬的细胞壁,这会阻碍细胞之间的融合。因此,为了实现两种不同植物体细胞的融合,首先需要去除细胞壁以制备原生质体(即去除了细胞壁的细胞)。常见的原生质体制备方法包括机械分离或酶解分离,其中酶解分离法使用更为广泛。
制备的原生质体可以通过物理或化学方法诱导其融合。诱导植物原生质体融合的物理方法中,使用比较普遍的是电融合法;化学方法中,使用最广泛的是用聚乙二醇(PEG)促融。此外,还可以使用盐类融合法、高钙离子浓度与高 pH 组合促融法、PEG 与高钙离子浓度和高 pH 组合促融法。
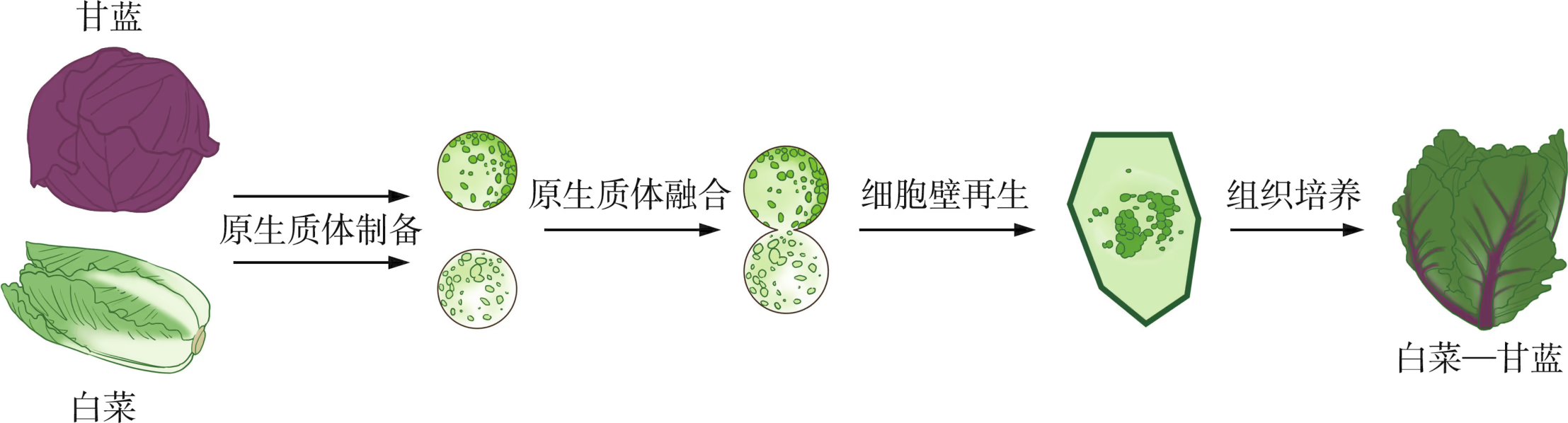
图 2-10 中箭头所示是一个成功融合后的杂合细胞,这一细胞既包含来自叶片细胞的叶绿体,也包含来自花瓣细胞的红色液泡。由于诱导操作对于细胞融合没有选择性且融合效率并不高,因此,除了形成不同类型的融合细胞外,还会存在大量未融合的细胞,这就需要使用一系列的方法将发生了融合的杂合细胞筛选出来,并对其进行细胞学和生化分析鉴定。
经筛选和鉴定过的融合细胞,可通过植物组织培养技术获得新植株。植物体细胞的杂交可以避开生殖细胞的受精过程,在亲缘关系较远的物种之间实现遗传物质的融合,从而创造出自然界中没有的新物种。但是也应当看到,大量的植物品种还没有建立起体细胞杂交的融合体系,并且要获得兼有双亲优良性状的杂合细胞还是非常困难的。此外,体细胞杂种植株的遗传稳定性和可育性也有待科研工作者的进一步研究。
动物细胞工程
动物细胞工程概述
动物细胞培养是指从动物体内取出组织或细胞,在体外模拟体内的生理条件,使之得以生存和生长并维持其结构与功能的技术。早在 1885 年,科学家就进行了动物组织离体培养的尝试。结果表明,离体后的动物组织或细胞在人工条件下仍然能够存活。真正意义上的现代动物细胞培养技术始于美国生物学家哈里森(R. G. Harrison)和法国科学家卡瑞尔(A. Carrel)的开创性工作。哈里森采用淋巴液作为培养基,成功培养了蛙胚神经组织。卡瑞尔特别注意到了实验中无菌操作的重要性,并发现鸡胚胎抽提液具有促进细胞生长的作用,成功培养了多种动物组织。随着对动物细胞离体生长机制的不断了解,人们开始广泛使用各种动物血清来促进细胞的生长,并开发出了人工合成的无血清培养基。同时,还设计出许多新型的动物细胞培养反应器,用于规模化培养。
动物细胞培养技术建立早期,直接采用包括血清在内的多种动物体液作为培养基。在研究和了解细胞生长所需成分的基础上,人们逐渐开发了一系列人工合成培养基。其中成分最简单的是低限量基础培养基(MEM),它包含了无机盐、微量元素、氨基酸、维生素和碳水化合物等 20 多种成分,但在使用时仍需要添加一定比例的动物血清,以补充细胞生长所必需的蛋白质、激素和生长因子等物质。
动物血清培养基固有的缺点主要是血清的成分复杂,会影响实验结果的分析;此外,不同批次的血清生产质量存在差异,会造成培养结果的不稳定。为了解决这些问题,科学家们致力于开发成分明确的无血清培养基。通过在基础培养基中添加相应的组分,以达到促进细胞的增殖和生长的要求。这些组分主要包括促进细胞贴壁的细胞外基质、各类生长因子和激素、酶抑制剂和某些蛋白质。这种成分明确的无血清培养基,已经成为细胞生物学基础研究领域的首选材料。
从动物体获取组织后,首先要通过机械法和消化法实现组织细胞的分散。消化法常用胰蛋白酶等试剂对动物组织块进行消化,使细胞从组织中分离出来。首次细胞培养称为原代培养。用于原代培养的动物活体组织往往由多种细胞组成,所以原代细胞培养物往往是多种细胞的混合物。原代培养后,将细胞从培养瓶中分离稀释,传到新的培养瓶中(传代培养),从而逐步分离获得单一类型的细胞。
在培养过程中,动物细胞非常容易受到各种微生物的污染,因此,必须要在严格无菌的环境下进行细胞培养,并在培养液中加入一定量的抗生素以防污染。同时,动物细胞培养对适宜的外界环境设定和维持也有严苛的要求。例如,适宜的培养温度(如哺乳动物细胞和禽类细胞的适宜温度为 )和 pH()是细胞在体外生长的必要条件;动物细胞的生长离不开氧气的供应,培养液中溶解氧水平会直接影响细胞的生长和代谢;通常在 的空气和 的 混合气体中培养,其中 气体是为保持培养液 pH 的稳定;还需要控制适宜的培养液渗透压;培养液中需包含维持动物细胞生长所必需的营养物质。此外,还要定期更换培养液,以便及时清除代谢产物,防止细胞代谢产物累积对细胞自身产生危害。
动物细胞工程常用的技术包括动物细胞培养、动物细胞融合和动物细胞核移植等,其中动物细胞培养是动物细胞工程的基础。人造皮肤的构建、动物分泌蛋白的规模化生产等,都离不开动物细胞培养。动物细胞培养是指从动物体中取出相关的组织,将它分散成单个细胞,然后在适宜的培养条件下,让这些细胞生长和增殖的技术。我们知道,动物体内的细胞之所以能够维持正常的生命活动,有些细胞还能不断增殖,是因为机体给这些细胞提供了适宜的条件,包括充足的营养、稳定的内环境等。在体外培养动物细胞,也需要满足类似的条件。
-
营养:细胞在体外培养时,培养基中应含有细胞所需要的各种营养物质。将细胞所需的营养物质按种类和所需量严格配制而成的培养基,称为合成培养基。由于人们对细胞所需的营养物质尚未全部研究清楚,因此在使用合成培养基时,通常需要加入血清等一些天然成分。培养动物细胞一般使用液体培养基,也称为培养液。
-
无菌、无毒的环境:在体外培养细胞时,必须保证环境是无菌、无毒的,即需要对培养液和所有培养用具进行灭菌处理以及在无菌环境下进行操作。培养液还需要定期更换,以便清除代谢物,防止细胞代谢物积累对细胞自身造成危害。
-
温度、 和渗透压:维持细胞生存必须有适宜的温度。哺乳动物细胞培养的温度多以 为宜。多数动物细胞生存的适宜 为 。此外,渗透压也是动物细胞培养过程中需要考虑的一个重要环境参数。
-
气体环境:动物细胞培养所需气体主要有 和 。 是细胞代谢所必需的, 的主要作用是维持培养液的 。在进行细胞培养时,通常采用培养皿或松盖培养瓶,并将它们置于含有 空气和 的 混合气体的 培养箱中进行培养。
动物细胞培养概述
进行动物细胞培养时,首先取健康动物的一小块组织并剪碎,用胰蛋白酶处理,使其分散成单个细胞,再与特定培养液混合,制成一定浓度的细胞悬液,最后转入培养瓶中,在适宜条件下培养。像这种直接从动物体获取组织或细胞后,进行的第一次培养过程称为原代培养,也叫初代培养。
在从动物体取出的成块组织中,细胞与细胞靠在一起,彼此限制了生长和增殖。因此,在进行细胞培养时,首先要对新鲜取材的动物组织进行处理,或用机械的方法,或用胰蛋白酶、胶原蛋白酶等处理一段时间,将组织分散成单个细胞。然后,用培养液将细胞制成细胞悬液,再将细胞悬液放入培养瓶或培养皿内,置于适宜环境中培养。体外培养的动物细胞可以分为两大类:一类细胞能够悬浮在培养液中生长增殖;另一类则需要贴附于某些基质表面才能生长增殖,大多数细胞属于这种类型,这类细胞往往贴附在培养瓶的瓶壁上,这种现象称为细胞贴壁。悬浮培养的细胞会因细胞密度过大、有害代谢物积累和培养液中营养物质缺乏等因素而分裂受阻。贴壁细胞在生长增殖时,除受上述因素的影响外,还会发生接触抑制现象,即当贴壁细胞分裂生长到表面相互接触时,细胞通常会停止分裂增殖。这时就需要对细胞进行分瓶培养,让细胞继续增殖。人们通常将分瓶之前的细胞培养,即动物组织经处理后的初次培养称为原代培养,将分瓶后的细胞培养称为传代培养。在进行传代培养时,悬浮培养的细胞直接用离心法收集;贴壁细胞需要重新用胰蛋白酶等处理,使之分散成单个细胞,然后再用离心法收集。之后,将收集的细胞制成细胞悬液,分瓶培养。
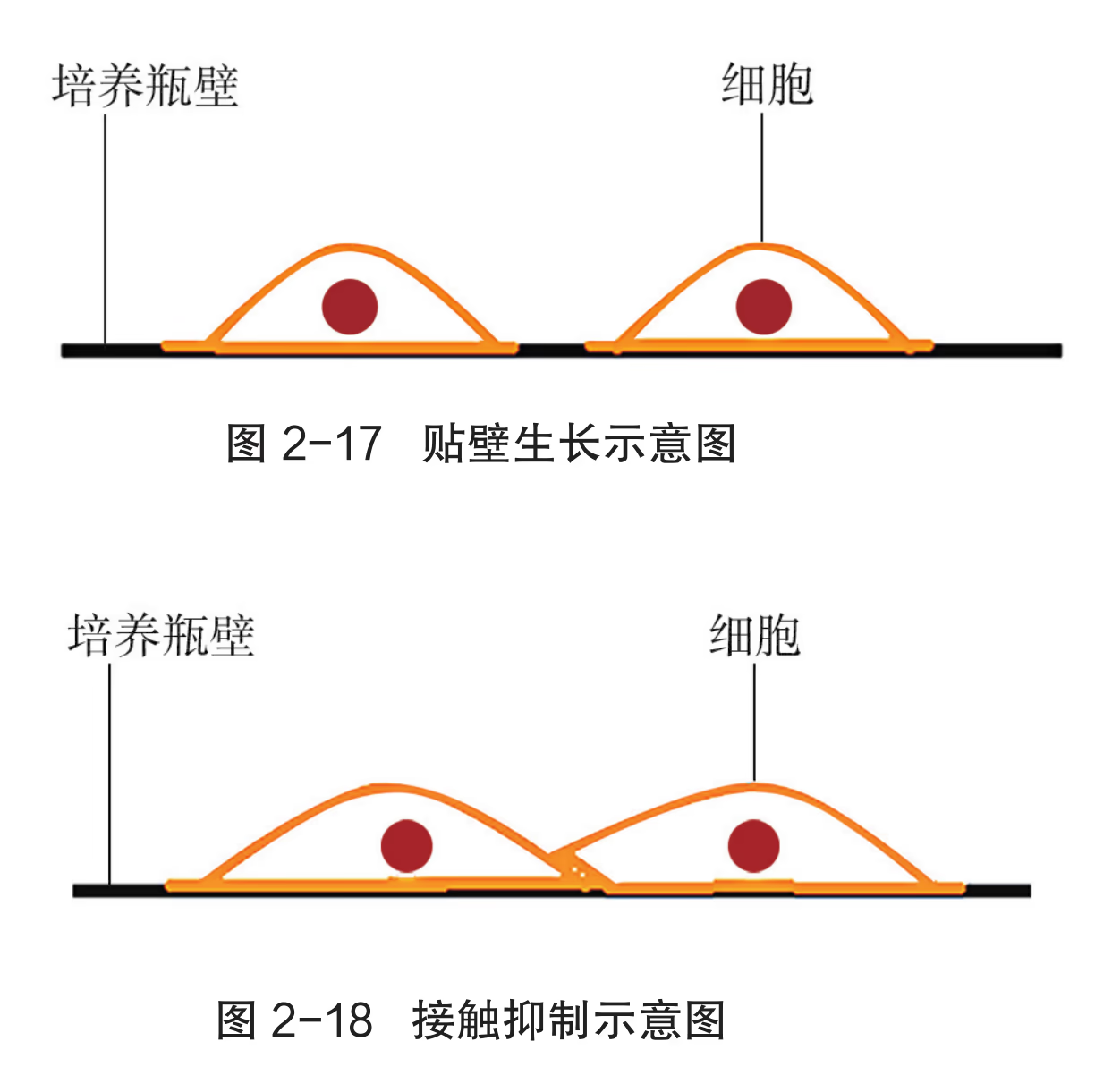
淋巴细胞、癌细胞等少部分细胞能够在悬浮状态下持续生长,而大部分动物细胞在培养液中悬浮生长一段时间后,会贴附在培养瓶的表面生长,这种现象称为贴壁生长。当培养的细胞生长到表面相互接触时,细胞就会停止分裂,逐渐走向衰亡,这种现象称为接触抑制。培养瓶中的细胞出现接触抑制后,需要用胰蛋白酶将细胞从瓶壁上脱离下来,重新分散稀释成细胞悬浮液,分装到多个培养瓶中继续培养。细胞由原代培养瓶中分离稀释后,转到新的培养瓶中继续培养的过程称为传代培养。
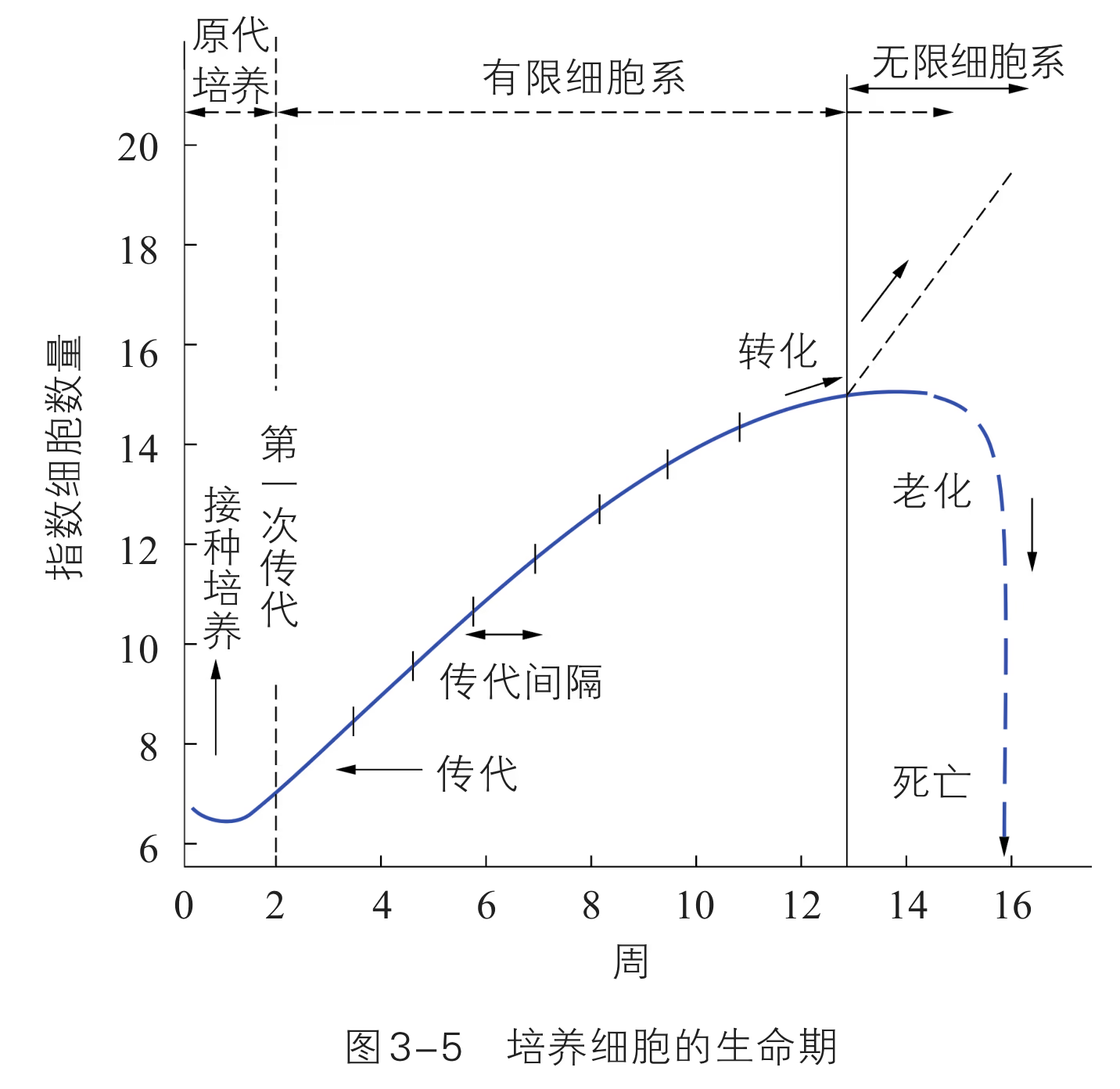
转瓶培养后的细胞一般传至 10 代左右,细胞的生长、分裂就会出现停滞,大部分细胞衰老死亡,但也有极少数细胞能存活下来,并可传至 40~50 代,这种传代细胞称为细胞株。一般情况下,当细胞株传至 50 代以后会再次出现停滞,不能继续传代,但有部分细胞的遗传物质发生了改变,并带有癌变的特点,从而有可能在适宜培养条件下无限传代下去,这种传代细胞叫作细胞系。
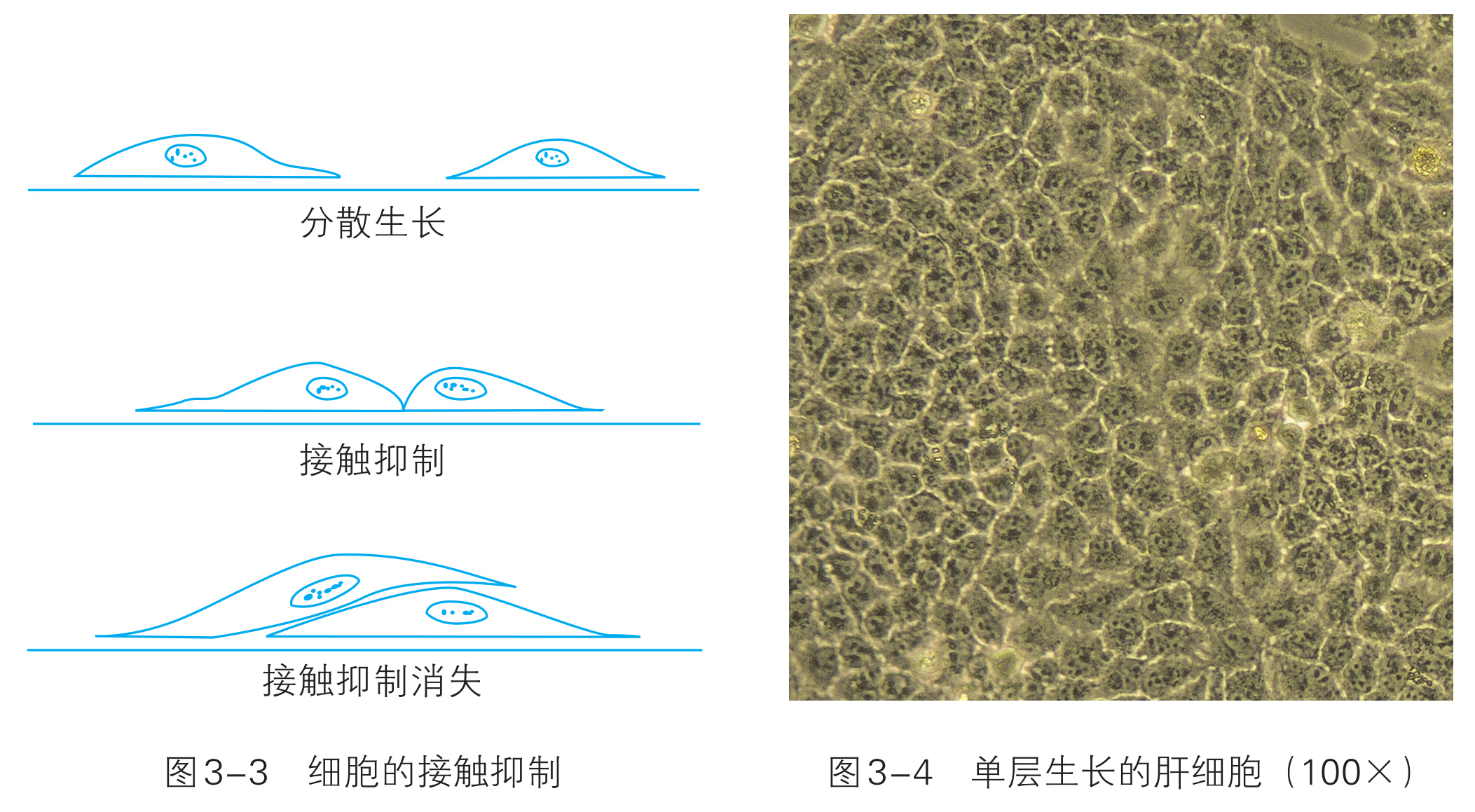
原代培养是指细胞或组织离开有机体后的首次培养。取材是原代培养的第一步,所取动物组织块一般体积大于 。若直接培养组织块,只有处于周边的少量细胞可以生存和繁殖,大部分中间细胞因不能获得穿透能力有限的营养物质而代谢不良。为了获取大量生长良好的细胞,必须先把组织细胞分散开,使细胞解离出来。 解离细胞的方法是先将组织块剪碎,用胰蛋白酶或胶原蛋白酶等消化处理一段时间,使组织松散,细胞分开。再将分散的细胞制成细胞悬液,进一步接种至培养瓶中,在人工控制的培养条件下,进行原代培养。 体外培养的动物细胞往往具有贴壁生长和接触抑制等特点。贴壁是有机体细胞在体内生存和生长发育的基本方式,包括细胞与细胞间的相互接触和细胞与细胞外基质之间的相互接触。正是基于细胞这种贴壁生长的特征,使得不同细胞之间结合而形成组织,也使得细胞和周围环境之间保持联系。体外培养时,绝大多数细胞仍然保持了贴壁生长的特性,必须贴附在某一固相支持物上如培养瓶壁上生长。来源于血液、淋巴组织的细胞以及某些肿瘤细胞不需要贴附在支持物上,而是呈悬浮生长状态。 接触抑制是体外细胞培养时贴壁型细胞生长的特性。一般情况下,正常的细胞在培养过程中不停顿地活动和运动,其外周的细胞膜呈现一些特征性的皱褶样活动。但是,当细胞由于移动而相互靠近发生接触时,细胞不再移动,接触区域的细胞膜皱褶样活动停止,最终细胞分裂停止,这种现象称为接触抑制(图 3-3)。由于细胞之间有接触抑制的特性,一般的正常细胞并不重叠于其他细胞之上生长,而是单层生长(图 3-4)。癌细胞由于无接触抑制而能够继续移动和增殖,导致细胞向三维空间扩展,发生堆积。因此,是否接触抑制可作为区别正常细胞与癌细胞的标志之一。
随着时间的延长,体外培养的细胞数量不断增加。当增长到一定程度后,由于发生接触抑制或培养空间的限制及营养物质的消耗枯竭,生长会逐渐减慢,甚至停止并死亡。因此,需要将原培养瓶内的细胞分离并稀释后转到多个培养瓶中,在新的培养基中进行传代培养。一般来说,原代培养的贴壁细胞达到生长基质的 表面面积后可进行传代培养。若传代过晚,可影响下一代细胞的机能状态。贴壁生长的细胞需经酶消化制成细胞悬液后才能传代。
原代培养物经首次传代成功后即成细胞系(cell line)。如图 3-5 所示,能连续培养下去的细胞系称为连续细胞系(continual cell line)或无限细胞系,不能连续培养下去的细胞系称为有限细胞系(finite cell line)。二倍体细胞通常为有限细胞系,如人的成纤维细胞可传 代,相当于 个细胞周期。连续细胞系被认为是发生转化了的细胞系(遗传物质发生改变),有的连续细胞系是恶性细胞系,有的连续细胞系虽已获得了不死性(immortality),但保留接触抑制现象,不致癌,是良性无限细胞系。
通过一定的选择或纯化方法,从原代培养物或细胞系中获得的具有特殊性质的细胞称为细胞株(cell strain)。细胞株一般具有恒定的染色体组型、病毒敏感性或抗性及其他生化特性。可以连续多次传代的细胞株称为连续细胞株(continual cell strain),可传代次数有限的细胞株称为有限细胞株(finite cell strain)。可见,细胞系是泛指可传代的细胞,细胞株是具有特殊性质的细胞系。 细胞株是细胞系经过进一步的克隆培养得到的。把一个单细胞从群体中分离出来单独培养,使之繁衍成一个新的细胞群体的技术,称为细胞克隆(cell cloning)或克隆培养(clone culture)。细胞克隆的最基本要求是:必须保证分离出来的细胞是一个而不是多个,即必须肯定所建成的克隆来源于单个细胞。经过克隆培养后的细胞群,由于来源于一个共同的祖细胞,基因型一致,遗传性状均一,是研究遗传规律和生理特性的重要材料。 细胞是构成生物有机体的基本结构和功能单位。在动物细胞、组织培养技术还不 成熟的时期,研究人员主要选取整个动物体或其组织、细胞予以固定,然后通过观察 来了解细胞的形态结构和生命活动。动物细胞培养技术完善之后,能直接观察到培养 细胞的动态变化过程,从而能直接观察、分析细胞的形态结构和真正意义上的生命活
动。培养正常或病变的细胞,可用于生理、病理、药理等方面的研究,例如,细胞全 能性的揭示、细胞周期调节控制的分析、癌变机理和衰老原因、抗癌药物的筛选、有 毒物质的检测等研究,都与细胞培养技术密不可分。 细胞培养在现实生活中具有重要意义,如皮肤细胞的培养用于烧伤患者的皮肤移 植,其他器官的细胞培养用于相关损伤组织器官的修复等。借助于某些细胞的大规模 培养获得细胞代谢产物,如获得动物疫苗、干扰素、单克隆抗体等有重要价值的生物 制品,是动物细胞培养的重要应用之一。
诱导性多能干细胞
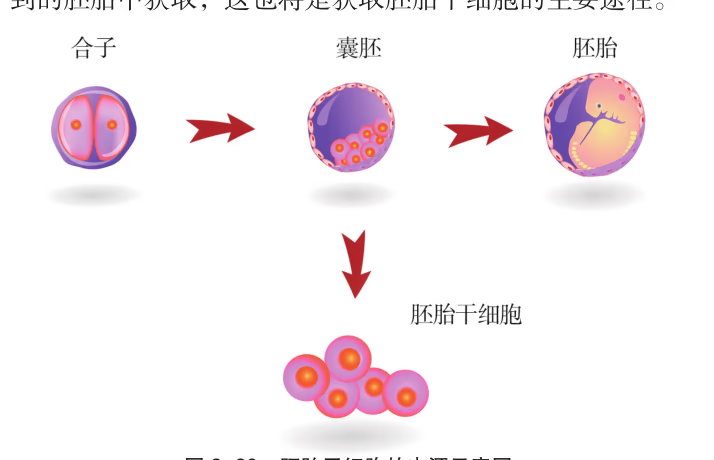
根据干细胞所处的发育阶段,可以把干细胞分成胚胎干细胞和成体干细胞。胚胎干细胞是高等动物受精卵发育成囊胚时的内细胞团,能在体外进行培养和传代,并在一定条件下保持分化的全能性,几乎可以分化出所有的细胞类型。胚胎干细胞特有的分化全能性以及可以无限传代增殖且不改变基因型和表现型的生物学特性,决定了它在生物学领域有着不可估量的应用价值。
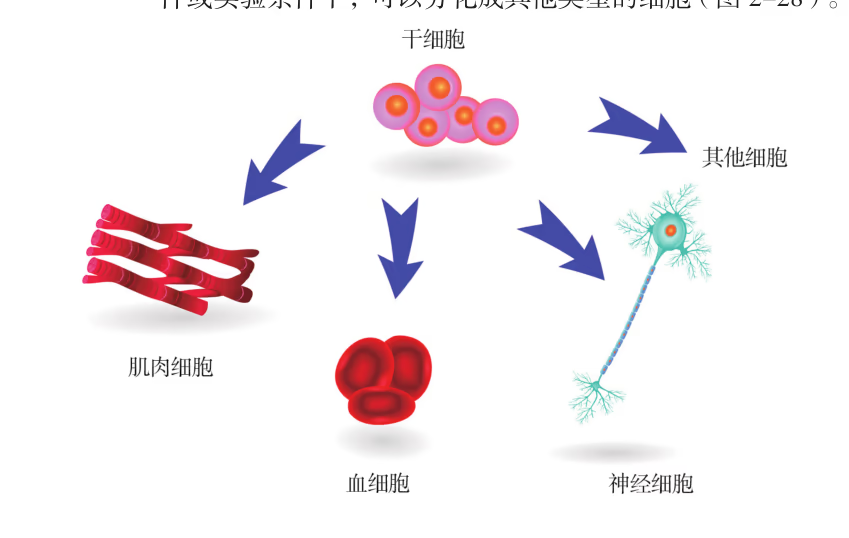
胚胎干细胞可取自囊胚的内细胞团或是胎儿的原始生殖细胞(图 2-22),其来源非常有限,而且它的获得和使用还涉及非常复杂的伦理问题。鉴于体细胞和干细胞拥有相同的遗传信息,目前可以通过多种技术手段将体细胞诱导形成具有与胚胎干细胞类似分裂和分化能力的干细胞,这样的细胞称为诱导性多能干细胞(iPS 细胞),为干细胞的广泛应用提供了基础。
在哺乳动物的克隆过程中,克隆后代的进一步发育还需要将早期胚胎移植入一个代孕母体的子宫内,我们将这种类型的克隆称为生殖性克隆。治疗性克隆的目的不是产生一个活体生物,而是产生胚胎干细胞,并将其在合适的条件下(如某些生长刺激型蛋白质存在)分化成各种特化细胞乃至形成组织或器官(图 2-23)。如果人们能找到正确的条件,便能培养胚胎干细胞用于修复受损或病变器官,如用来置换因脊髓损伤或心肌梗死而损坏的细胞。
干细胞的培养成功是动物细胞培养领域重大的成就之一,在一定条件下,干细胞可以分化成其他类型的细胞。干细胞存在于早期胚胎、骨髓和脐带血等多种组织和器官中,包括胚胎干细胞和成体干细胞等。成体干细胞是分布在成体组织中尚未分化的、具有自我更新潜能的干细胞,在组织或系统的修复和再生中起着关键作用(图 2-24)。
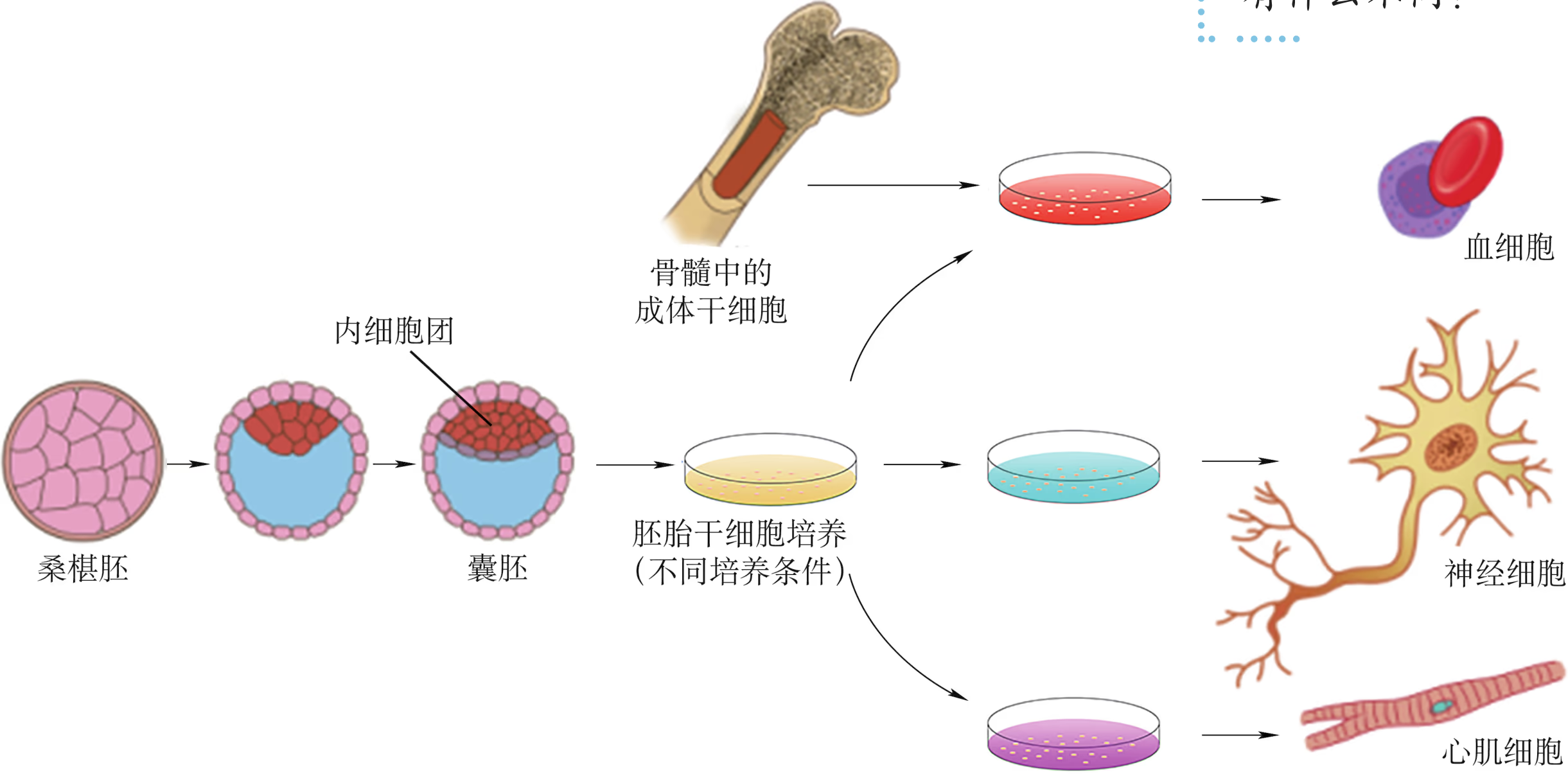
按照分化潜能的大小,可以将成体干细胞分成两种类型。一类是多能干细胞,具有分化出多种细胞组织的能力,但不具备发育成完整个体的能力,如骨髓多能造血干细胞,可以分化出多种不同的血细胞(图 2-25)。另一类是单能干细胞,只能分化成一种类型或密切相关的两种类型的细胞,如上皮组织基底层的干细胞、肌肉中的成肌细胞等。
胚胎干细胞(简称 ES 细胞)存在于早期胚胎中,具有分化为成年动物体内的任何一种类型的细胞,并进一步形成机体的所有组织和器官甚至个体的潜能,ES 细胞可以在体外分化成心肌细胞、神经元和造血干细胞等细胞。成体干细胞是成体组织或器官内的干细胞,包括骨髓中的造血干细胞、神经系统中的神经干细胞和睾丸中的精原干细胞等。一般认为,成体干细胞具有组织特异性,只能分化成特定的细胞或组织,不具有发育成完整个体的能力。造血干细胞是发现最早、研究最多、应用也最为成熟的一类成体干细胞,主要存在于成体的骨髓、外周血和脐带血中。
有着自我更新能力及分化潜能的干细胞,与组织、器官的发育、再生和修复等密切相关,因而在医学上有着广泛的应用。例如,将正常的造血干细胞移植到病人体内,恢复病人的造血和免疫功能,已成为治疗白血病及一些恶性肿瘤放疗或化疗后引起的造血系统、免疫系统功能障碍等疾病的一种重要手段;神经干细胞在治疗神经组织损伤和神经系统退行性疾病(如帕金森病、阿尔茨海默病等)方面有重要的应用价值。
胚胎干细胞必须从胚胎中获取,这涉及伦理问题,因而限制了它在医学上的应用。2006 年,科学家通过体外诱导小鼠成纤维细胞,获得了类似胚胎干细胞的一种细胞,将它称为诱导多能干细胞(简称 iPS 细胞),并用 iPS 细胞治疗了小鼠的镰状细胞贫血。现在,用 iPS 细胞治疗阿尔茨海默病、心血管疾病等领域的研究也取得了新进展。因为诱导过程无须破坏胚胎,而且 iPS 细胞可以来源于病人自身的体细胞,将它移植回病人体内后,理论上可以避免免疫排斥反应,所以科学家普遍认为 iPS 细胞的应用前景优于胚胎干细胞。
科学家已尝试采用多种方法来制备 iPS 细胞,包括借助载体将特定基因导入细胞中,直接将特定蛋白导入细胞中或者用小分子化合物等来诱导形成 iPS 细胞。iPS 细胞最初是由成纤维细胞转化而来的,后来发现已分化的 T 细胞、B 细胞等也能被诱导为 iPS 细胞。干细胞的研究正在如火如荼地开展。虽然将干细胞用于临床治疗还面临一些问题,如存在导致肿瘤发生的风险,但是我们相信,随着理论和技术的不断完善,干细胞将在再生医学、药物安全性与有效性检测等领域发挥大作用。
早在 20 世纪 50 年代,人们就开始使用骨髓移植的方法治疗血液系统疾病。到 80 年代末,外周血干细胞移植技术开始逐渐替代传统的骨髓移植。随后,又在脐带血中发现了脐血干细胞。与骨髓和外周血干细胞移植相比,脐血干细胞移植具有无来源限制、对人类白细胞抗原(human leukocyte antigen,简称 HLA,是决定移植后排斥的主要因素)配型要求较低、不易受到病毒或肿瘤的污染等优点。我国已经掌握了脐血干细胞分离、纯化、冷冻保存和复苏的全套技术,并在上海建立了我国第一个脐血库(图 2-26)。
此外,胰岛干细胞最初是从尚未发病的糖尿病小鼠胰导管中分离得到的。将胰岛干细胞经体外诱导分化成产胰岛素的胰岛 细胞并移植给糖尿病小鼠,接受移植的小鼠血糖浓度控制良好,这为干细胞治疗糖尿病奠定了基础。皮肤干细胞是另一种成体干细胞。我国科学家发现,人体烧伤皮肤原位处存在着皮肤干细胞。在一定的药物诱导下,这样的皮肤干细胞能使烧伤处的皮肤原位再生。这是我国在皮肤干细胞研究及临床上的重大突破。
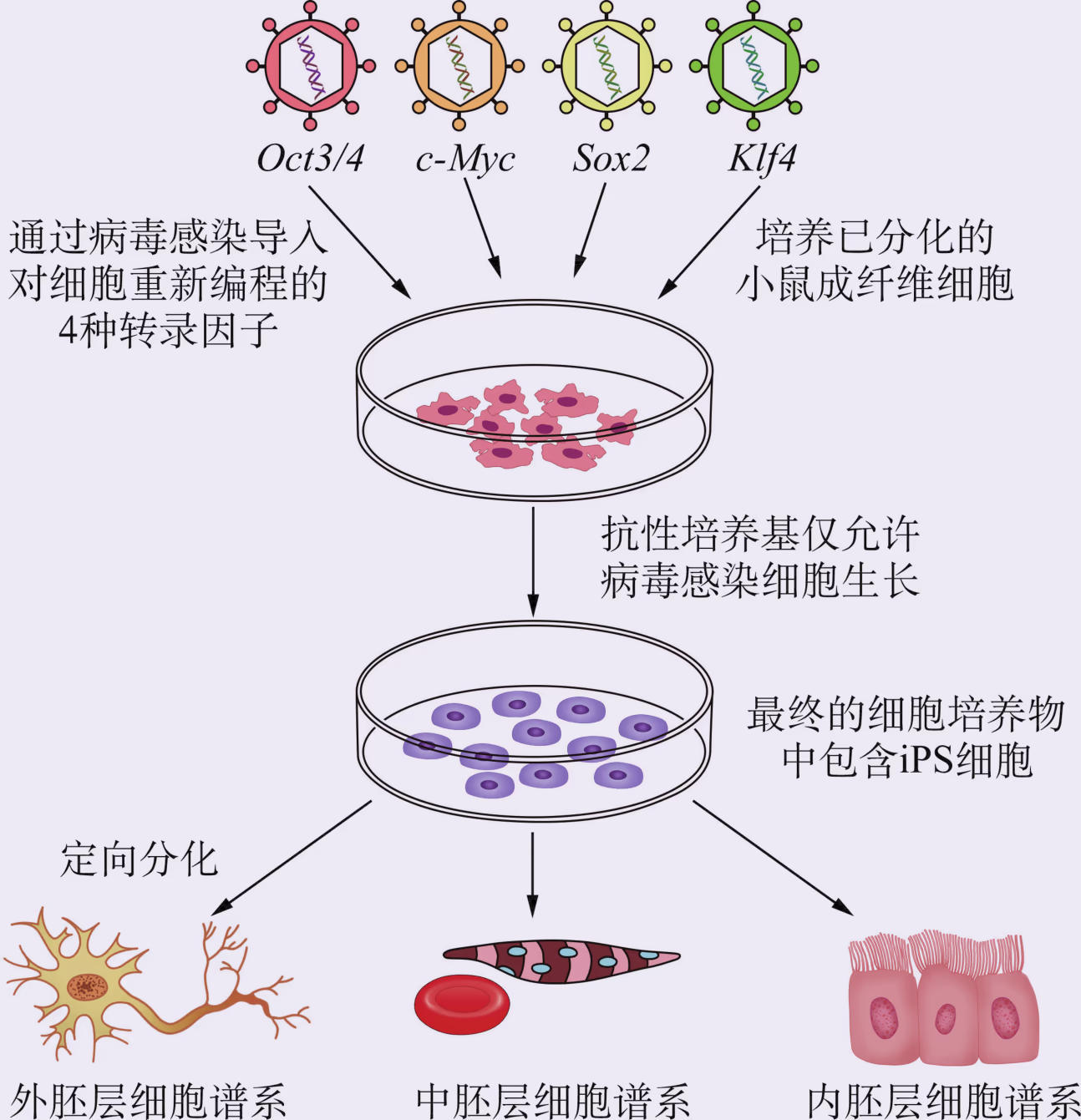
胚胎干细胞具有分化全能性,但其来源有限且涉及复杂的伦理问题;而各种成体干细胞(除造血干细胞外)的采集往往在技术上非常困难,且不具备分化成所有细胞类型的能力。这些因素都限制了干细胞在临床上的应用。由于体细胞与干细胞都携带着相同的遗传信息,因此,只要能对体细胞的遗传信息表达进行重新编程,理论上就能够获得具有与干细胞类似分化能力的细胞。日本科学家山中伸弥(S. Yamanaka)于 2006 年首先报道了将 4 种转录因子基因组合转入分化的体细胞中,使其重编程,最终成功获得了类似胚胎干细胞的一种细胞类型,即诱导性多能干细胞(图 2-27)。山中伸弥因此获得了 2012 年的诺贝尔生理学或医学奖。随后,世界各国科学家陆续发现其他方法同样也可以制造这种细胞。
iPS 细胞技术的出现,在干细胞生物学、表观遗传学以及再生医学研究领域都引起了强烈的反响。在基础研究方面,利用 iPS 细胞作为实验模型,可大大加速对多能性调控机理的深入研究。在实际应用方面,iPS 细胞的获得方法相对简单和稳定,不需要使用卵细胞或者胚胎,这在技术上和伦理上都比其他方法更有优势。iPS 细胞的建立进一步拉近了干细胞与临床疾病治疗的距离,在细胞替代性治疗以及发病机理的研究、新药筛选方面具有巨大的潜在应用价值。我国科学家在 iPS 细胞研究领域取得了一系列重大成果。2009 年,研究团队从小鼠皮肤细胞培育出世界首例来自 iPS 细胞的可育小鼠,证实了 iPS 细胞发育全能性这一关键科学问题。2022 年,我国科学家在国际上首次实现了运用化学小分子将人成体细胞转变为 iPS 细胞。这些重大原创性成果的取得,使我国在干细胞基础理论和应用的一些领域中处于国际引领地位。
组织工程是干细胞应用研究的重要方向之一。组织工程的基本原理是:首先从自体或异体组织中分离干细胞,经体外扩增达到一定的细胞数量后,将这些细胞作为种子细胞种植在预先构建好的聚合物骨架上。在适当的生长条件下,细胞沿着聚合物骨架生长和分化,最终发育成具有特定形态及功能的工程组织。借助组织工程技术,科学家们已经获得了组织工程化的皮肤、骨骼、血管等器官,有望解决临床上急需的人工组织与器官问题(图 2-28)。
1975 年,科学家建立了淋巴细胞杂交瘤技术,解决了单克隆抗体制备的难题(图 2-14)。在该流程中,从众多细胞中筛选出能产生针对某一特定抗原决定簇的杂交瘤细胞,是单克隆抗体制备的关键。科学家采用缺失次黄嘌呤-鸟嘌呤磷酸核糖转移酶(HGPRT)基因的骨髓瘤细胞与经绵羊红细胞免疫的小鼠脾脏细胞进行融合,然后将经融合操作后的细胞置于添加了含有次黄嘌呤(H)、氨基蝶呤(A)和胸腺嘧啶(T)的动物细胞培养基(HAT 培养基)中进行筛选,成功得到了杂交瘤细胞。已知氨基蝶呤能阻断次黄嘌呤核苷酸和胸腺嘧啶核苷酸的生物合成(图 2-32)。
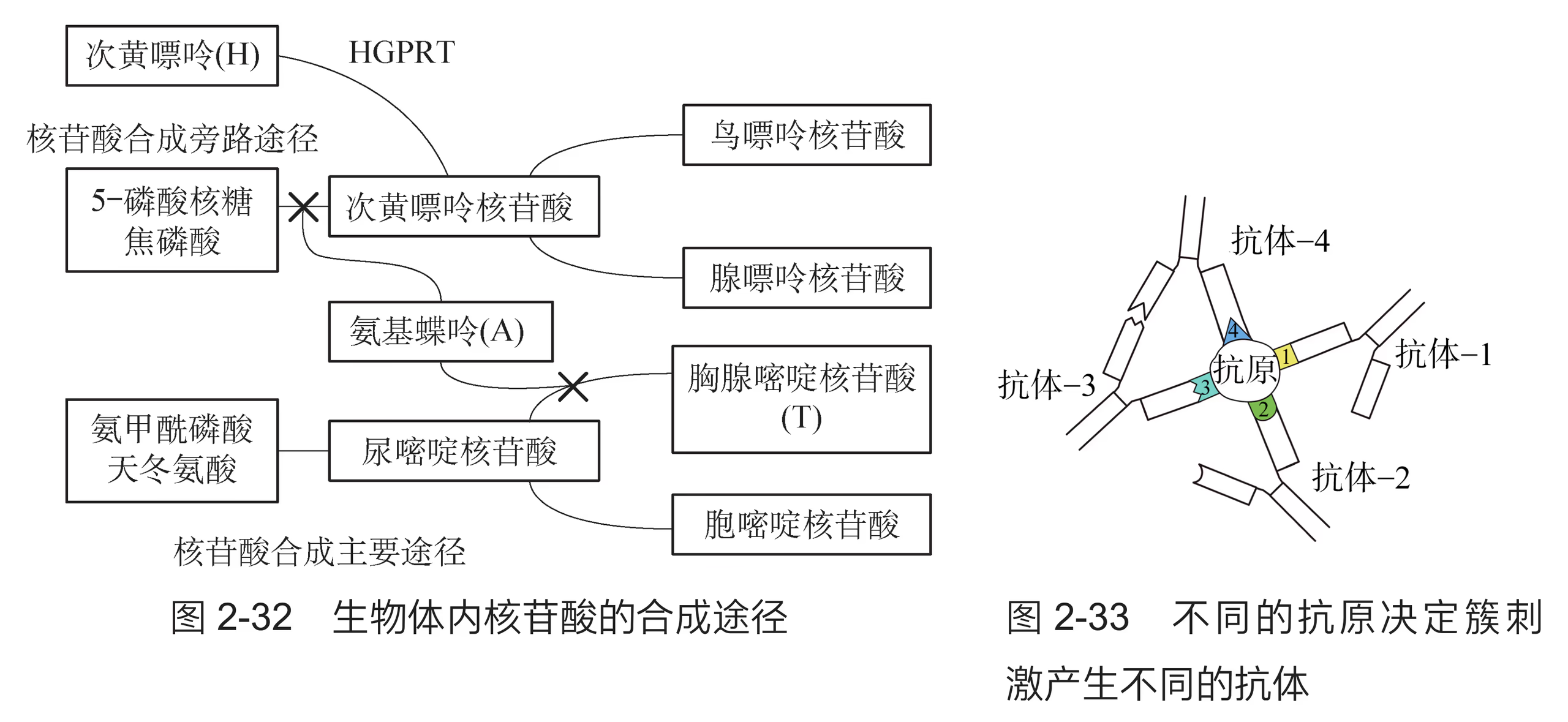
经过 HAT 培养基筛选出的杂交瘤细胞仍有多种类型,每一种杂交瘤细胞产生的抗体特异性针对某个抗原决定簇(图 2-33)。因此,需要经过第二次筛选才能获得针对某一抗原决定簇的特异性 B 淋巴细胞。第二次筛选通常采用稀释法,即将杂交瘤细胞多倍稀释,接种在多孔的细胞培养板上,使每一孔含一个或几个杂交瘤细胞,而后进行相应的处理以便筛选出所需的单个杂交瘤细胞。
动物细胞融合技术
受精过程是生殖细胞之间的自然融合。此外,科学家陆续发现体细胞之间也会有融合现象:首先是在脊椎动物肿瘤细胞中观察到了多核现象;随后,在骨髓、肺等正常组织及炎症或坏死部位也都发现了多核现象(图 2-13)。细胞培养技术问世后,体细胞融合在体外培养的肿瘤细胞中得到了验证。现代生物工程中的细胞融合指的是体细胞杂交,即在离体条件下,用人工方法将不同生物或同种生物不同类型的单细胞融合成一个杂合细胞。
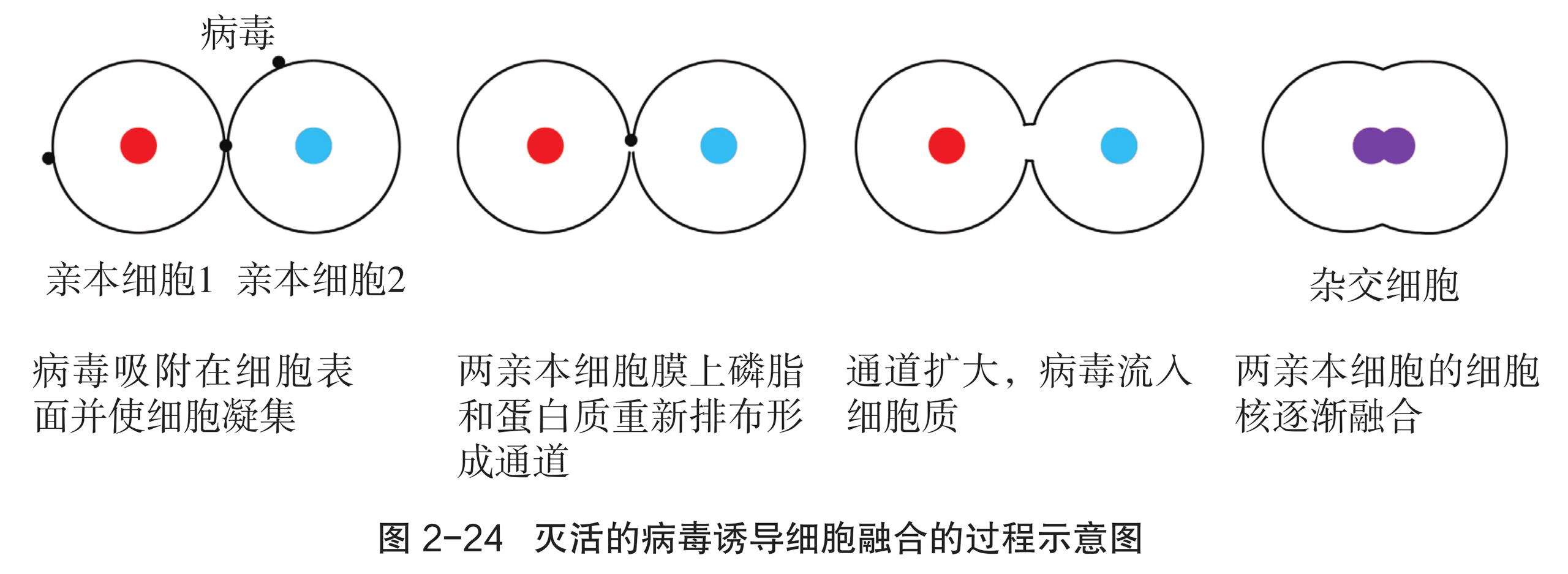
某些病毒(灭活)如仙台病毒可以诱发细胞融合形成多核细胞,这一发现开创了细胞人工融合的新领域。除了病毒促融外,细胞融合还可采用物理、化学等手段,使两个甚至多个动物细胞融合形成一个细胞。物理方法如电融合,在高压脉冲电场的作用下,细胞膜表面会形成大量微膜孔,进而发生细胞融合现象。化学方法如聚乙二醇(PEG)或其衍生物,具有比仙台病毒高数百倍的促细胞融合效率。细胞融合技术可以使人们按照预先的设计使不同的细胞融合,创造出新的杂合细胞。
聚乙二醇诱导融合法因其使用方便且诱导细胞融合的成功率高,是较为常用的细胞融合方法。当然,聚乙二醇也有一定的局限性,如有一定毒性,对某些细胞(如卵细胞)不适用。聚乙二醇分子因能改变细胞质膜的结构,而被广泛地作为促进细胞融合的化学试剂,用于促进形成杂种细胞。
::: {.grid} { width="100%" } { width="60%" } :::
单克隆抗体问世后,因其特异性强而被大量应用于临床。单克隆抗体既可以用于疾病的诊断,也可用于疾病的治疗。如利用单克隆抗体可特异性识别抗原物质的特点,研制出了具有精确"定位"能力的抗癌"药物导弹",它能将药物准确地引导到癌细胞表面并予以攻击,以提高抗癌药物的特异性和抗癌效率。
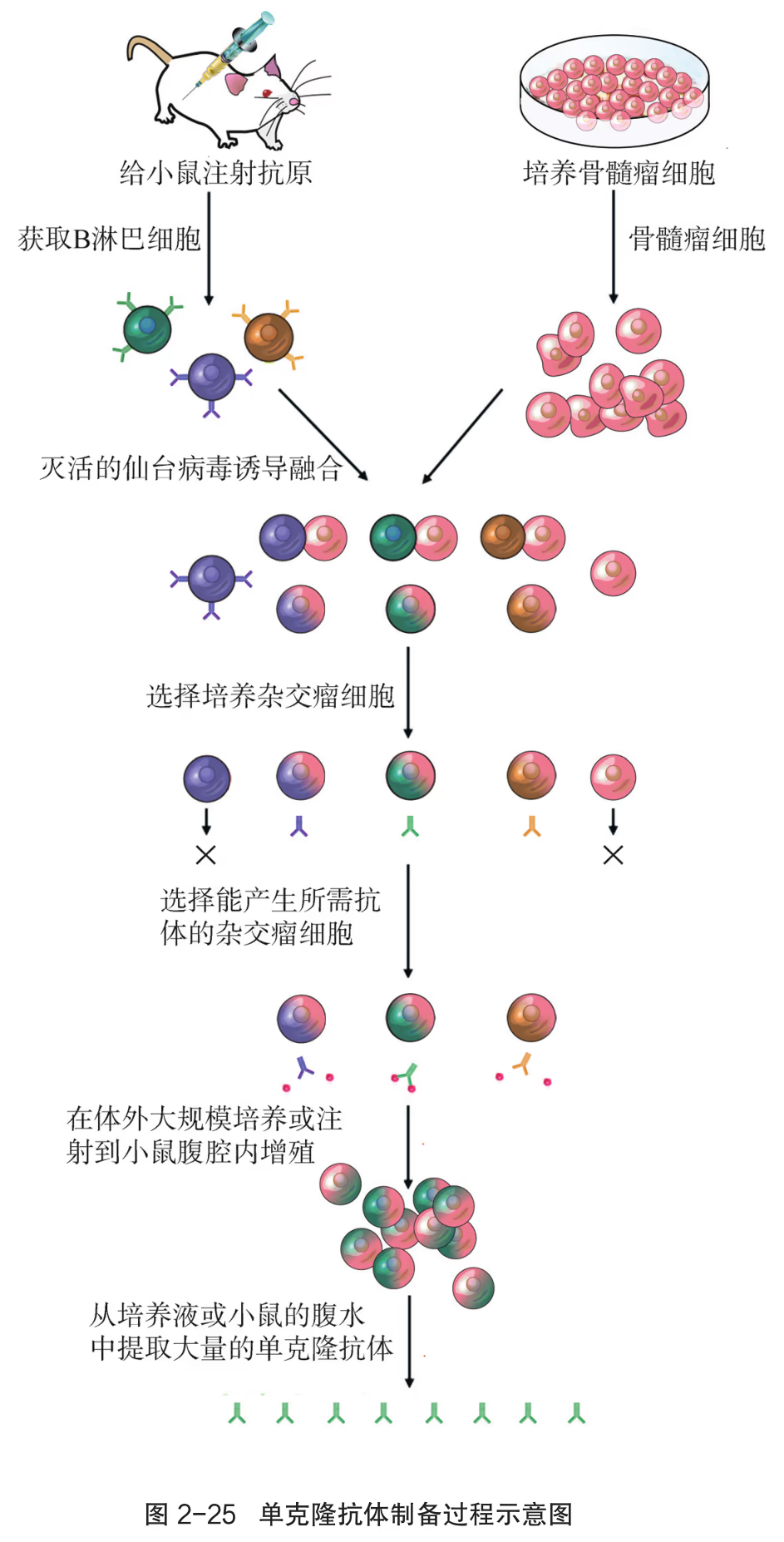
动物细胞融合技术比较突出的成就是单克隆抗体的制备。抗体是机体免疫应答的一个重要组成部分,可以特异性识别抗原。但绝大部分的抗原拥有多个不同的抗原决定簇(即抗原分子中决定抗原特异性并刺激 B 淋巴细胞分化为浆细胞的特定区域),会同时刺激多个不同的 B 淋巴细胞形成克隆群。所以,针对同一个抗原,机体往往会产生多种抗体。有些抗体通常存在特异性不强、质量不稳定等问题,且经抗原激活后的 B 淋巴细胞也无法在体外长时间培养。那么,是否可以得到针对某一特定抗原决定簇的、高度均一的抗体呢?科学家们在患骨髓瘤的小鼠体内发现了骨髓瘤细胞,该细胞在体外培养时能无限增殖。于是,科学家便设想:是否可以利用细胞融合技术,将 B 淋巴细胞的单一抗体合成和分泌能力与骨髓瘤细胞的体外无限增殖特性融为一体呢?
为了验证这一设想,1975 年,德国科学家克勒(G. Köhler)和阿根廷科学家米尔斯坦(C. Milstein)以绵羊红细胞作为抗原刺激小鼠脾脏的 B 淋巴细胞,获得了能产生抗体的浆细胞,后者在促融剂的作用下与小鼠骨髓瘤细胞发生融合,形成了杂交瘤细胞。通过选择培养基筛选出抗体分泌阳性的杂交瘤细胞,并对其进行克隆培养(图 2-14)。这样获得的杂交瘤细胞既能无限增殖,又能持续分泌高度均一、仅针对某一特定抗原决定簇的抗体,即单克隆抗体。
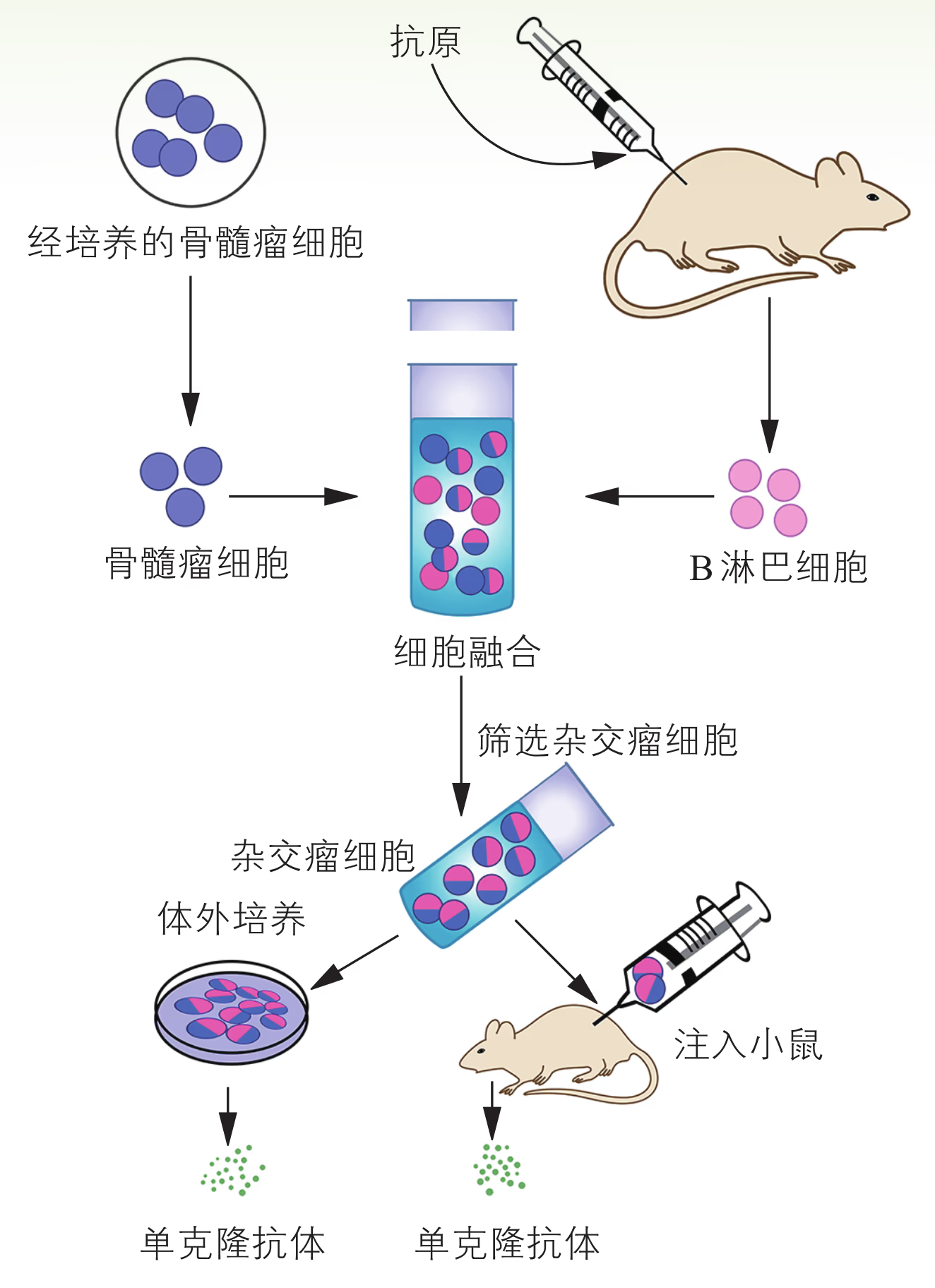
制备单克隆抗体的流程主要有:抗原准备及动物免疫、细胞融合、杂交瘤细胞的筛选、杂交瘤细胞的增殖及抗体提取等。单克隆抗体的高度特异性可用于鉴定各种抗原,对多种疾病的诊断、治疗以及生理过程的检测具有重要价值。例如,利用单克隆抗体的靶向作用,可将针对某一肿瘤抗原的单克隆抗体与化疗药物或放疗物质连接,将其携带至靶器官,直接杀伤靶细胞,降低对健康组织与细胞的伤害(即靶向性治疗)。再如,早孕检测试纸可通过尿液快速检测出受孕妇女胎盘中产生的人绒毛膜促性腺激素。
动物细胞融合技术就是使两个或多个动物细胞结合形成一个细胞的技术。融合后形成的杂交细胞具有原来两个或多个细胞的遗传信息。动物细胞融合与植物原生质体融合的基本原理相同。诱导动物细胞融合的常用方法有 PEG 融合法、电融合法和灭活病毒诱导法等。灭活是指用物理或化学手段使病毒或细菌失去感染能力,但并不破坏它们的抗原结构。灭活病毒诱导细胞融合的原理是:病毒表面含有的糖蛋白和一些酶能够与细胞膜上的糖蛋白发生作用,使细胞互相凝聚,细胞膜上的蛋白质分子和脂质分子重新排布,细胞膜打开,细胞发生融合。细胞融合技术突破了有性杂交的局限,使远缘杂交成为可能。人们可以按照预先的设计使不同细胞融合,至今,种间、属间、科间,甚至动物和植物之间的细胞融合都已获得了成功。目前,这一技术已经成为研究细胞遗传、细胞免疫、肿瘤和培育生物新品种等的重要手段,特别是利用动物细胞融合技术发展起来的杂交瘤技术,为制造单克隆抗体开辟了新途径。
早期人们为了获得抗体,就向动物体内反复注射某种抗原,使动物产生抗体,然后从动物血清中分离所需抗体。用这种方法制备的抗体不仅产量低、纯度低,而且特异性差。为了解决这一难题,科学家进行了多年的研究和探索。他们发现,哺乳动物感染病原体后,体内会形成相应的 B 淋巴细胞,这些细胞能分泌抗体,抗体识别并特异性结合病原体,从而抑制病原体的增殖等。动物体内产生的特异性抗体的种类超过百万种,但每一个 B 淋巴细胞只分泌一种特异性抗体。因此,要想获得大量的单一抗体,必须克隆单一的 B 淋巴细胞,形成细胞群。1975 年,英国科学家米尔斯坦和德国科学家科勒在前人工作的基础上,充分发挥想象力,设计了一个极富创造性的实验方案。他们想到,如果把一种 B 淋巴细胞与能在体外大量增殖的骨髓瘤细胞融合,所得到的融合细胞就可能大量增殖,产生足够数量的特定抗体。根据该设想,通过实验,他们得到了单克隆抗体。
-
用特定的抗原对小鼠进行免疫,并从该小鼠的脾中得到能产生特定抗体的 B 淋巴细胞。
-
用特定的选择培养基进行筛选:在该培养基上,未融合的亲本细胞和融合的具有同种核的细胞都会死亡,只有融合的杂交瘤细胞才能生长。
-
对上述经选择培养的杂交瘤细胞进行克隆化培养和抗体检测,经多次筛选,就可获得足够数量的能分泌所需抗体的细胞。
-
将抗体检测呈阳性的杂交瘤细胞在体外条件下大规模培养,或注射到小鼠腹腔内增殖。
-
从细胞培养液或小鼠腹水中获取大量的单克隆抗体。
使用高效的细胞毒素类药物进行化疗可以有效杀伤肿瘤细胞。但细胞毒素没有特异性,在杀伤肿瘤细胞的同时还会对健康细胞造成伤害,这限制了它在临床上的应用。抗体---药物偶联物通过将细胞毒素与能特异性识别肿瘤抗原的单克隆抗体结合,实现了对肿瘤细胞的选择性杀伤。ADC 通常由抗体、接头和药物(如细胞毒素)三部分组成,它的作用机制如下图所示。
单克隆抗体能准确地识别抗原的细微差异,与特定抗原发生特异性结合,并且可以大量制备,因此被广泛用作诊断试剂,在多种疾病的诊断和病原体鉴定中发挥重要的作用。例如,利用同位素或荧光标记的单克隆抗体在特定组织中成像的技术,可定位诊断肿瘤、心血管畸形等疾病。除前面介绍的单克隆抗体可以运载药物外,单克隆抗体自身也能用于治疗疾病。在医药领域,单克隆抗体药物占有非常重要的地位。
动物细胞核移植技术
细胞核移植是将一种动物细胞的细胞核移植至去核的卵母细胞中,并使重组细胞发育成新胚胎,继而发育成动物个体的过程。所产生的后代具有与细胞核供体基本相同的遗传成分。科学家首先用两栖类动物成功地尝试了此技术,并陆续在绵羊、小鼠、牛、山羊等哺乳动物中获得了胚胎细胞核或体细胞核移植的后代。
细胞核移植技术是目前动物克隆的主要手段。所谓克隆(clone)是指分子、细胞或个体的无性繁殖系,如从一个分子复制成一组分子称为分子克隆;从一个细胞分裂生成一群细胞称为细胞克隆;动植物个体的无性繁殖就是个体克隆。
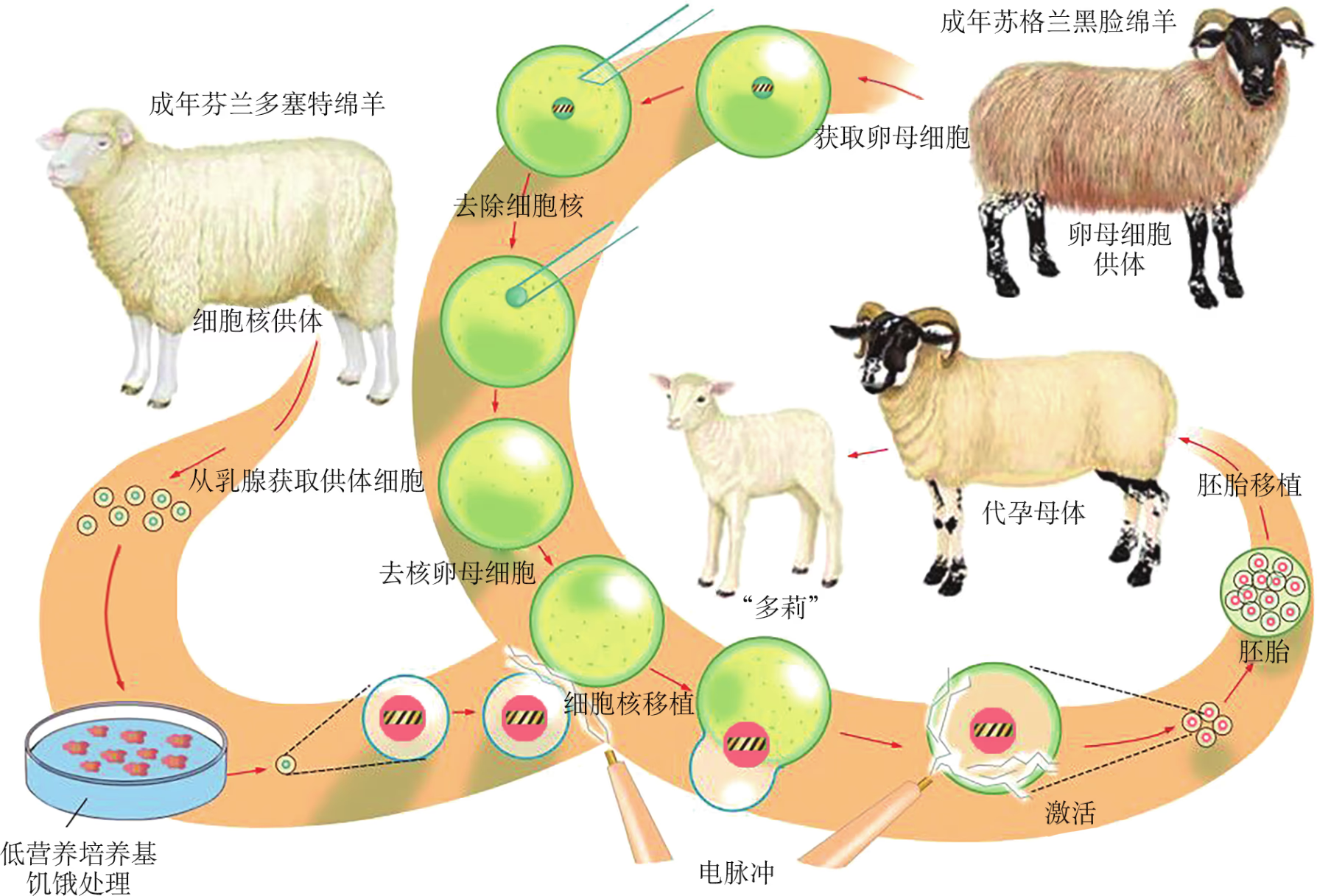
动物克隆技术通过显微操作,将供体细胞的核移入去核的卵母细胞中(图 2-16),经过人工活化和体外培养后,再移植入代孕母体内,使其发育成为含有与供体细胞相同遗传物质的个体,该过程便是典型的个体克隆。克隆羊"多莉"是人类首次用成年动物的体细胞细胞核移植获得的克隆后代(图 2-17)。"多莉"的诞生证明了哺乳动物高度分化的体细胞核能够在卵母细胞细胞质的作用下恢复其全能性,为细胞全能性理论增添了有力的实验证据。
采用体细胞克隆技术成功获得了克隆羊、克隆小鼠和克隆牛后,科学家又向与人类相近的灵长类动物的体细胞克隆发起了挑战。2017 年,在中国上海诞生了世界首例非人灵长类动物体细胞克隆后代"中中"和"华华"(图 2-18)。体细胞克隆猴的成功以及未来基于体细胞克隆猴的疾病模型的创建,将有效缩短药物研发周期、提高药物研发成功率。这将使我国率先发展出基于非人灵长类动物疾病模型的全新医药研发产业链,促进针对阿尔茨海默病、自闭症等脑疾病以及人类免疫缺陷、肿瘤、代谢性疾病的新药研发进程。
克隆技术具有广泛的应用前景。在农牧业中,可将具有特殊期望性状的动物通过克隆产生更多的畜群;在科学研究中,基因完全相同的动物可为有关实验提供最佳的"对照动物";在制药行业中,人们正在尝试用克隆动物生产具有潜在医疗用途的产品;在野生物种保护中,可利用克隆技术繁殖濒危物种,为生态保护带来了无限希望(图 2-19)。然而,克隆技术本身还有很多问题未能解决。不断增加的证据显示,克隆动物的健康水平较差,很多克隆动物表现出诸如肥胖、肺炎、肝功能低下、早衰性死亡等方面的缺陷。例如,克隆绵羊"多莉"寿命仅 6 岁,因身患通常仅在年老绵羊中才会出现的肺部疾病并发症,于 2003 年死亡,而与其共同饲养的非克隆同伴预期寿命可达 12 年。
动物细胞核移植技术是将动物一个细胞的细胞核移入去核的卵母细胞中,使这个重新组合的细胞发育成新胚胎,继而发育成动物个体的技术。哺乳动物核移植可以分为胚胎细胞核移植和体细胞核移植。由于动物胚胎细胞核分化程度低,表现全能性相对容易,而动物体细胞分化程度高,表现全能性十分困难,因此动物体细胞核移植的难度明显高于胚胎细胞核移植。而在动物体细胞核移植中,非人灵长类动物的体细胞核移植非常困难。主要原因一方面是供体细胞的细胞核在去核卵母细胞中不能完全恢复其分化前的功能状态,这导致了胚胎发育率低;另一方面是对非人灵长类动物胚胎进行操作的技术尚不完善。
体细胞核移植技术在畜牧业、医药卫生以及其他领域有着广泛的应用前景。在畜牧生产中,利用体细胞核移植技术,可以加速家畜遗传改良进程、促进优良畜群繁育。在医药卫生领域,通过建立转基因体细胞系,再利用体细胞核移植技术培育的转基因克隆动物可以作为生物反应器,生产许多珍贵的医用蛋白;在治疗人类疾病时,转基因克隆动物的细胞、组织或器官可以用于异种移植;以患者作为供体培育的人核移植胚胎干细胞,经过诱导分化能形成相应的细胞、组织或器官,将它们移植给患者时可以避免发生免疫排斥反应。此外,研究克隆动物可使人类更深入地了解胚胎发育及衰老过程;克隆一批遗传背景相同的动物,可以通过它们之间的对比来分析致病基因;克隆特定疾病模型的动物,还能为研究该疾病的致病机制和开发相应的药物提供帮助。在保护濒危物种方面,该技术有望增加濒危物种的存活数量。
尽管通过体细胞核移植技术已经获得了多种克隆动物,但该技术的成功率仍然非常低,各个技术环节也有待进一步改进。相对于技术研究,核移植的理论研究较为滞后,需要与发育生物学、细胞生物学和分子遗传学等学科更深层次的理论研究相结合。此外,一些研究者指出,绝大多数克隆动物存在健康问题,许多克隆动物表现出遗传和生理缺陷,如体形过大、异常肥胖、发育困难、脏器缺陷和免疫失调等。总之,体细胞核移植技术的研究仍在继续深入,人类对克隆技术的应用还有许多问题需要解决。期待在不远的将来,克隆技术能更好地造福于人类。
基因工程简述
基因工程是在生物化学、分子生物学和微生物学等学科的基础上发展起来的,正是这些学科的基础理论和相关技术的发展催生了基因工程。基因工程是指按照人们的愿望,通过转基因等技术,赋予生物新的遗传特性,创造出更符合人们需要的新的生物类型和生物产品。从技术操作层面看,由于基因工程是在 DNA 分子水平上进行设计和施工的,因此又叫作重组 DNA 技术。
-
1944 年,艾弗里(O. Avery, )等通过肺炎链球菌的转化实验,不仅证明了遗传物质是 DNA,还证明了 DNA 可以在同种生物的不同个体之间转移。
-
1950 年,埃德曼(P. Edman, )发明了一种测定氨基酸序列的方法。2 年后,桑格(F. Sanger, )首次完成了对胰岛素氨基酸序列的测定。
-
1953 年,沃森(J. D. Watson, )和克里克(F. Crick, )建立了 DNA 双螺旋结构模型并提出了遗传物质自我复制的假说。
-
1958 年,梅塞尔森(M. Meselson, )和斯塔尔(F. W. Stahl, )用实验证明了 DNA 的半保留复制。几乎同一时期确立的中心法则阐明了遗传信息流动的方向。
-
1961 年,尼伦伯格(M. W. Nirenberg, )和马太(J. H. Matthaei, )破译了第一个编码氨基酸的密码子。截至 1966 年,64 个密码子均被成功破译。
-
1967 年,科学家发现,在细菌拟核 DNA 之外的质粒具有自我复制能力,并可以在细菌细胞间转移。
-
1970 年,科学家在细菌中发现了第一个限制性内切核酸酶(简称限制酶)。
-
20 世纪 70 年代初,多种限制酶、DNA 连接酶和逆转录酶相继被发现。这些发现为 DNA 的切割、连接以及功能基因的获得创造了条件。
-
1972 年,伯格(P. Berg, )首先在体外进行了 DNA 改造的研究,成功地构建了第一个体外重组 DNA 分子。
-
1973 年,科学家证明质粒可以作为基因工程的载体,构建重组 DNA,导入受体细胞,使外源基因在原核细胞中成功表达,并实现物种间的基因交流。至此,基因工程正式问世。
-
1977 年,桑格等科学家发明了 DNA 序列分析的方法,为基因序列图的绘制提供了可能。此后,DNA 合成的问世为体外合成 DNA 提供了方便。
-
1982 年,第一个基因工程药物------重组人胰岛素被批准上市。基因工程药物成为世界各国研究和投资开发的热点。
-
1983 年,科学家采用农杆菌转化法培育出世界上第一例转基因烟草。此后,基因工程进入了迅速发展的阶段。
-
1984 年,我国科学家朱作言()领导的团队培育出世界上第一条转基因鱼。
-
1985 年,穆里斯(K. Mullis, )等人发明了 PCR,为获取目的基因提供了有效手段。
-
1990 年,人类基因组计划启动。2003 年,该计划的测序任务顺利完成。
-
2013 年,华人科学家张锋()及其团队首次报道利用最新的基因组编辑技术------CRISPR(成簇规律间隔短回文重复)技术编辑了哺乳动物基因组。该技术可以实现对特定基因的定点插入、敲除或替换。
-
21 世纪以来,科学家发明了多种高通量测序技术,可以实现低成本测定大量核酸序列,加速了人们对基因组序列的了解。
上述仅是按照时间顺序简要提及基因工程相关基础理论的突破和技术的创新。有些你已经学习过,更多的将在本章中展开。科学提供对世界的说明,技术将科学原理应用于认识自然、造福人类的实践,工程则综合运用多种技术来生产人类需要的产品。科学、技术、工程和社会的互动,不断调整着人类与自然界的关系,推动着文明的进步。
基因工程的基本原理
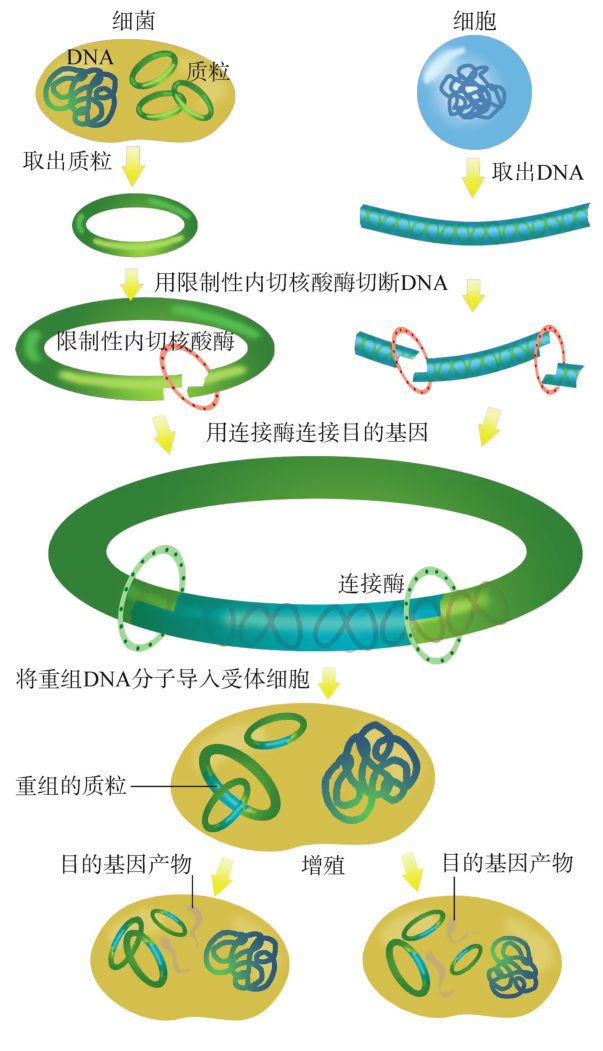
基因工程是指将一种或多种生物(供体)的基因与运载工具在体外进行拼接重组,然后转入另一种生物体(受体)内,使之按照人们的意愿遗传并表达出新产物或新性状。由于重组拼接的基因和运载工具都是 DNA 分子,因此,基因工程也称为重组 DNA 技术。对于有性生殖的生物而言,不同物种之间存在生殖隔离,难以进行基因交流;即便生活在同一个栖息地的大多数细菌物种也很难交换遗传信息。重组 DNA 技术诞生的科学意义在于打破这种隔离,使跨物种间基因的定向转移成为可能。另一方面,发酵工程所用的微生物菌种以及细胞工程所涉及的细胞株,很多都是采用基因工程技术改造过的,因此基因工程是现代生物工程的核心技术。
对于更加详细的基因工程的技术方面,请查阅基因工程及其技术苏教版基础资料.pdf。
1973 年,基因工程在遗传学、微生物学、生物化学和分子生物学等生物科学分支学科的基础上问世。20 世纪 70 年代初,科学家考恩和博耶等人首先尝试将两种不同来源的 DNA 片段(鼠伤寒沙门氏菌的 RSF1010 和大肠杆菌的 pSC101)在试管里进行拼接,然后将这种重组 DNA 分子导入大肠杆菌细胞中,结果发现这种人工组装的 DNA 分子不仅能准确复制自己,而且还能表达两种 DNA 的遗传信息。紧接着,他们又将另一种亲缘关系较远的金黄色葡萄球菌基因成功导入大肠杆菌中,同样观察到该"外源"基因在大肠杆菌中表达出原来的遗传性状。对上述实验结果进行归纳与概括,他们认为不同物种的细菌可以在实验条件下毫无障碍地进行"交流" 。然而,这一结论是否也能演绎和外推至亲缘关系更远的真核生物物种中呢?随后,他们又参照前人建立的实验程序,成功地将非洲爪蟾的一些基因植入大肠杆菌并获得成功表达,从而证实了他们先前提出的假说:遗传物质 DNA 是可以在保持其功能的前提下进行人为拼接的。自此,宣告了具有划时代里程碑意义的基因工程的诞生。
理论基础:
-
揭示 DNA 是遗传物质:1944 年,著名的肺炎双球菌转化实验不仅揭示了生物体的遗传物质是 DNA,同时还显示 DNA 可以从一种生物个体转移到另一种生物个体。这项遗传学研究是基因工程中基因转移操作的先驱性工作。
-
确立 DNA 双螺旋结构和中心法则:1953 年,DNA 双螺旋模型的建立促进了 DNA 半保留复制的实验证明。随后不久确立的中心法则解开了 DNA 复制、转录和 mRNA 翻译过程之谜,阐明了遗传信息传递的方向。这些分子生物学原理为基因工程中提升目的基因的复制和表达水平奠定了基础。
-
破译遗传密码:1963~1966 年期间,遗传密码的破译不仅使人们认识到自然界几乎所有生物共用一套遗传密码,而且为基因的鉴别和合成等提供了理论依据。
技术支撑:
-
发现运载工具:1953 年,发现细菌细胞内除了拟核 DNA 外,还存在一类具有独立复制能力的小型 DNA(称为质粒),它们可在细菌细胞之间转移,这一发现为基因转移找到了一种运载工具。
-
开发工具酶:1970 年,在细菌中鉴定了第一种能切割DNA 的酶;随后,又相继分离纯化了多种能连接 DNA 和具有其他功能的酶。这些生物化学发现为 DNA 的切割、连接以及基因的获取奠定了技术基础。
-
实现 DNA 体外重组:1972 年,首次在体外尝试 DNA 的切割和拼接操作,并成功地构建了第一个体外重组 DNA 分子。
基于上述理论和技术的建立,1973 年,美国科学家考恩(S. N. Cohen)和博耶(H. W. Boyer)等人在体外构建出含有四环素和青霉素两个抗性基因的重组 DNA 分子,将之导入大肠杆菌后,这种重组 DNA 分子能稳定复制,并使大肠杆菌表达出相应的抗生素抗性,由此宣告了基因工程的诞生。考恩在评价其实验结果时指出,基因工程技术完全有可能使大肠杆菌具备其他生物物种的特殊生物功能,如光合作用和抗生素合成等。
微生物以其无性繁殖、生长迅速以及易于基因操作等优势为基因工程的问世作出了重大贡献。每个经基因工程改造过的细胞就如同性能优良的微型生物反应器,可用来高效生产人们感兴趣的目标产物。然而,如何控制这些微型生物反应器的数量以及单个微型生物反应器的生产性能,还需要运用生化工程学的理论和技术。
基因工程自 20 世纪 70 年代诞生后,经历了飞速发展,已成为生命科学和生物工程的核心技术。基因工程在医学、农牧业、食品工业等众多领域有着广泛的应用。
基因治疗:就单基因突变或缺陷所导致的疾病而言,向患者体内输入正常版本的基因至少在理论上有可能矫正遗传病,这便是人体基因治疗的基本策略。1990 年,美国批准了世界首例人体基因治疗的临床研究计划,对一名因腺苷脱氨酶基因(ADA)缺陷而患有重度联合免疫缺陷病(SCID)的儿童(终身只能生活在保护性的"气泡"中,被称为"泡泡儿")进行基因治疗。这一项目最终获得成功,从而开创了基因治疗的新纪元。临床医学研究人员定期从患者血液中分离免疫细胞,用携带正常 ADA 等位基因的病毒表达载体感染这些免疫细胞,然后将这些得以矫正的细胞回输至患者体内,即所谓的"间接体内疗法"。
然而,这种治疗方案在给 SCID 患者带来福音的同时,却也暴露出严重的副作用:四名接受治疗的患者后来发展为白血病,其中一名患者因治疗基因的插入激活了癌基因而死亡。基因治疗给人们带来了希望,但很少有证据能显示其应用的安全性和有效性。尽管如此,临床和学术界仍在坚持不懈地探究更新更严格的安全治疗策略。
2017 年,美国食品和药品管理局(FDA)终于批准了该国的首例基因治疗用剂,该用剂是一种受体蛋白经蛋白质工程设计和改造过的患者自体 T 细胞,用于治疗某些儿童和青少年的急性淋巴细胞白血病(ALL)。生理性 T 细胞受体(TCR)只有在受到抗原刺激后才能发挥免疫效应,而肿瘤表面抗原较难激活 T 细胞。用特异性识别特定肿瘤表面抗原的单链抗体(scFv)取代 TCR 的 αβ 链,同时对 TCR 复合物进行结构改造,构成所谓的嵌合型抗原受体(CAR)。与生理性 T 细胞受体(TCR)复合物相比,第二代嵌合型抗原受体(CAR)在结构上发生了很大改变。
将这种携带工程化 CAR 的 T 细胞作为治疗用剂输回患者体内,此类疗法统称为 CAR-T 细胞疗法。在体内,这种工程化 CAR 能指导 T 细胞识别并杀死表面上带有相应抗原的肿瘤细胞。然而,科学研究没有止境,人们担心这种第二代 CAR-T 细胞输回患者体内后是否会形成新的免疫原性。综上所述,以首例基因治疗临床试验(补充或更换 ADA 基因)为代表的众多策略均属于忠实于原基因编码序列的基因工程;而以 CAR-T 细胞疗法(修饰或改造受体蛋白)为代表的新型策略则属于第二代基因工程------蛋白质工程。
基因工程在农牧业方面的应用
基因工程在农牧业中的应用发展迅速。 年,全世界转基因作物的种植面积增加了一百多倍,从 发展到 (图 3-9)。2016 年世界范围的统计数据表明,转基因作物的种植使化学杀虫剂施用量减少了 ,作物产量增加了 ,增加经济收益近 1.3 万亿元。美国是世界上转基因作物种植面积最大的国家,转基因棉花、大豆、玉米的种植面积占相关作物种植面积的比例都超过了 。2017 年,我国转基因作物的种植面积位居世界第八位,商业化种植的转基因作物有棉花和番木瓜。
在转基因动物方面,近些年几乎每年都有令人瞩目的研究成果报道,有些成果正在进入实用化和商业化开发的阶段。2015 年 11 月,第一种用于食用的转基因动物------转基因大西洋鲑(俗称"三文鱼")在美国获得批准上市。
目前,基因工程技术已被广泛用于改良动植物品种、提高作物和畜产品的产量等方面。
-
转基因抗虫植物:从某些生物中分离出具有抗虫功能的基因,将它导入作物中培育出具有抗虫性的作物,是目前防治作物虫害的一种发展趋势。已问世的转基因抗虫植物有转基因抗虫棉花、玉米、大豆、水稻(图 3-10)和马铃薯等。
-
转基因抗病植物:许多栽培作物由于自身缺少抗病基因,因此用常规育种的方法很难培育出抗病的新品种。科学家将来源于某些病毒、真菌等的抗病基因导入植物中,培育出了转基因抗病植物,如转基因抗病毒甜椒(图 3-11)、番木瓜和烟草等。
-
转基因抗除草剂植物:杂草常常危害农业生产,而大多数除草剂不仅能杀死田间杂草,还会损伤作物,导致作物减产。将降解或抵抗某种除草剂的基因导入作物,可以培育出抗除草剂的作物品种。这样在喷洒除草剂时,田间杂草会被杀死而作物不会受到损伤。目前已经获得转基因抗除草剂玉米(图 3-12)、大豆(图 3-13)、油菜和甜菜等。
-
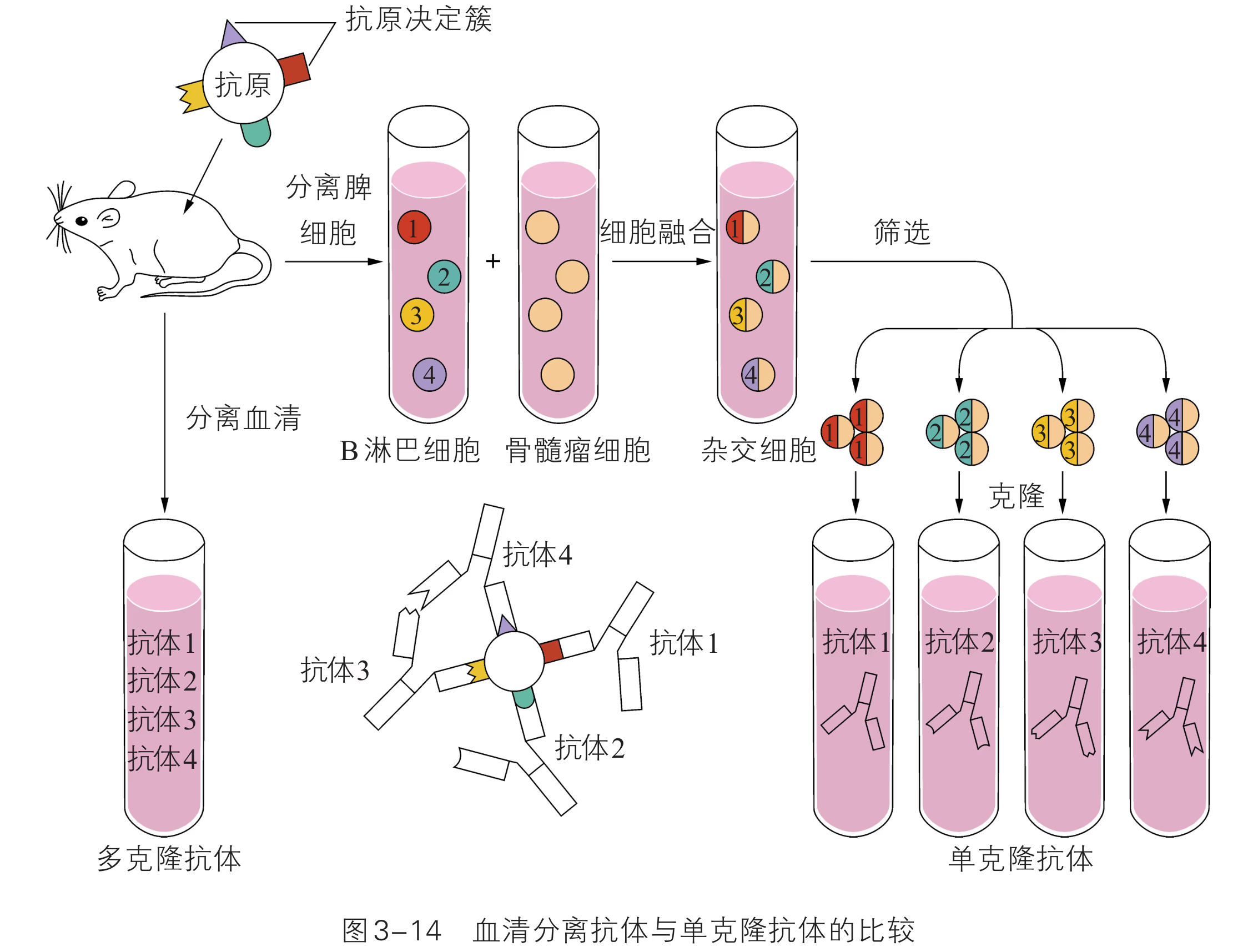
改良植物的品质:随着生活水平的提高,人们越来越关注植物的营养价值、观赏价值等,利用转基因技术可以改良这些品质。例如,将某种必需氨基酸含量多的蛋白质编码基因导入植物中,可以提高这种氨基酸的含量,科学家培育的某种转基因玉米中赖氨酸的含量比对照高 。我国科学家成功地将与植物花青素代谢相关的基因导入矮牵牛中,使它呈现出自然界没有的颜色变异,大大提高了它的观赏价值(图 3-14)。
-
提高动物的生长速率:由于外源生长激素基因的表达可以使转基因动物生长得更快,因此科学家将这类基因导入动物体内,以提高动物的生长速率。例如,我国科学家将外源生长激素基因导入鲤鱼,在同等养殖条件下,转基因鲤鱼的生长速率比非转基因鲤鱼提高了 (图 3-15)。
-
改善畜产品的品质:有些人由于乳糖酶分泌少,不能完全消化牛奶中的乳糖,食用牛奶后会出现腹泻等不适症状,这称为乳糖不耐受。我国约有 的成年人对乳糖不耐受。为了解决这一问题,科学家将肠乳糖酶基因导入奶牛基因组,使获得的转基因牛分泌的乳汁中,乳糖的含量大大降低,而其他营养成分不受影响。
基因工程在医药卫生领域的应用
对微生物或动植物的细胞进行基因改造,使它们能够生产药物,是目前基因工程取得实际应用成果非常多的领域。这些药物包括细胞因子、抗体、疫苗和激素等,它们可以用来预防和治疗人类肿瘤、心血管疾病、传染病、糖尿病和类风湿关节炎等。我国生产的重组人干扰素、血小板生成素、促红细胞生成素和粒细胞集落刺激因子等基因工程药物均已投放市场(图 3-16)。
干扰素是一种具有干扰病毒复制作用的糖蛋白,在临床上被广泛用于治疗病毒感染性疾病。此外,干扰素对治疗乳腺癌、淋巴瘤、多发性骨髓瘤和某些白血病等也有一定的疗效。传统生产干扰素的方法是从人血液中的白细胞内提取,每 血液只能提取出 干扰素。 年,科学家用基因工程方法从大肠杆菌及酵母菌细胞内获得了干扰素,从 培养物中可以得到 干扰素。1993 年,我国批准生产重组人干扰素 -1b,它是我国批准生产的第一个基因工程药物,目前主要用于治疗慢性乙型肝炎、丙型肝炎等。
利用基因工程技术,还可以让哺乳动物批量生产药物。科学家将药用蛋白基因与乳腺中特异表达的基因的启动子等调控元件重组在一起,通过显微注射的方法导入哺乳动物的受精卵中,由这个受精卵发育成的转基因动物在进入泌乳期后,可以通过分泌乳汁来生产所需要的药物,这称为乳腺生物反应器或乳房生物反应器。目前,科学家已经在牛、山羊等动物乳腺生物反应器中,获得了抗凝血酶、血清白蛋白、生长激素和 -抗胰蛋白酶等重要的医药产品。
基因工程技术还可能使建立移植器官工厂的设想成为现实。目前,人体移植器官短缺是一个世界性难题。为此,人们不得不把目光移向寻求可替代的移植器官。由于猪的内脏构造、大小、血管分布与人的极为相似,而且与灵长类动物相比,猪体内隐藏的、可导致人类疾病的病毒要少得多,是否可以用猪的器官来解决人类器官移植的来源问题呢?实现这一目标的最大难题是免疫排斥。目前,科学家正尝试利用基因工程技术改造猪的器官,采用的方法是在器官供体的基因组中导入调控基因表达的 DNA 序列,以抑制抗原决定基因的表达,或设法除去抗原决定基因,再结合克隆技术,培育出不会引起免疫排斥反应的转基因克隆猪器官。
1981 年,采用人血培养病毒制得的乙肝疫苗率先在美国批准上市。这是人类历史上第一种商业化的乙肝疫苗,象征着人类对抗乙肝的一次革命性突破。但是,有限的来源和高昂的价格使其难以普及,而且存在通过血液感染其他病毒(如艾滋病病毒)的风险也引起了人们的严重担忧。基因工程技术为乙肝疫苗的生产工艺带来了革命性突破:首先,获得乙肝病毒表面蛋白(用作抗原)的基因,然后将其转移至酵母中,可使酵母高效合成乙肝病毒表面抗原蛋白,用作乙肝疫苗。由于酵母易于大规模发酵培养,且整个生产过程能彻底摆脱血液需求和病毒培养,因而通过基因工程大规模生产乙肝疫苗成为了可能。
自 20 世纪 80 年代以来,基因工程开始朝着高等动植物物种的遗传性状改良以及人体基因治疗等方向发展。1982 年,科学家将大鼠的生长激素基因转入小鼠体内,培育出具有大鼠雄健体魄的转基因小鼠。1990 年,美国首次批准了一项人体基因治疗临床研究计划,对一名因腺苷脱氨酶基因缺陷而患有重度联合免疫缺陷病的儿童进行基因治疗并获得成功,从而开创了基因治疗的新纪元。
基因工程在食品工业方面的应用
利用基因工程菌除了可以生产药物,还能生产食品工业用酶、氨基酸和维生素等。例如,阿斯巴甜是一种普遍使用的甜味剂,主要由天冬氨酸和苯丙氨酸形成,这两种氨基酸就可以通过基因工程实现大规模生产。
奶酪是一种被广泛食用的发酵奶制品。大多数奶酪的生产需要使用凝乳酶来凝聚固化奶中的蛋白质。传统的制备凝乳酶的方法是通过杀死未断奶的小牛,然后将它的第四胃的黏膜取出来进行提取。随着人口的增长和人们饮食需求的变化,单纯依靠宰杀小牛来获得凝乳酶的方法已不能满足日益增长的市场需求。于是,科学家将编码牛凝乳酶的基因导入大肠杆菌、黑曲霉或酵母菌的基因组中,再通过工业发酵批量生产凝乳酶。用这种方法生产的凝乳酶已于 1990 年投入市场,截至 2016 年,有超过 17 个国家在用它生产奶酪。
加工转化糖浆需要的淀粉酶,加工烘烤食品要用到的脂肪酶等也都可以通过构建基因工程菌,然后用发酵技术大量生产。相比从天然产物中提取的酶,用基因工程技术获得的工业用酶的纯度更高,而且它的生产成本显著降低,生产效率较高。
基因工程使人们更容易培育出具有优良性状的动植物品种,获得很多过去难以得到的生物制品,甚至还能培育出可以降解多种污染物的"超级细菌"来处理环境污染,利用经过基因改造的微生物来生产能源...... 未来,期待基因工程带给我们更多的惊喜。
DNA 片段的扩增及电泳鉴定
利用 PCR 可以在体外进行 DNA 片段的扩增。PCR 利用了 DNA 的热变性原理,通过调节温度来控制 DNA 双链的解聚与结合。PCR 仪实质上就是一台能够自动调控温度的仪器。一次 PCR 一般要经历 30 次循环。
DNA 分子具有可解离的基团,在一定 下,这些基团可以带上正电荷或负电荷。在电场的作用下,这些带电分子会向着与它所带电荷相反的电极移动,这个过程就是电泳。PCR 的产物一般通过琼脂糖凝胶电泳来鉴定。在凝胶中 DNA 分子的迁移速率与凝胶的浓度、DNA 分子的大小和构象等有关。凝胶中的 DNA 分子通过染色,可以在波长为 的紫外灯下被检测出来。
-
用微量移液器按照右栏中的配方或 PCR 试剂盒的说明书,在微量离心管中依次加入各组分。
-
待所有的组分都加入后,盖严离心管的盖子。将微量离心管放入离心机里,离心约 ,使反应液集中在管的底部。
-
参照下表的参数,设置好 PCR 仪的循环程序。将装有反应液的微量离心管放入 PCR 仪中进行反应。
-
根据待分离 DNA 片段的大小,用电泳缓冲液配制琼脂糖溶液,一般配制质量分数为 的琼脂糖溶液。在沸水浴或微波炉内加热至琼脂糖熔化。稍冷却后,加入适量的核酸染料混匀。
-
将温热的琼脂糖溶液迅速倒入模具,并插入合适大小的梳子,以形成加样孔。
-
待凝胶溶液完全凝固后,小心拔出梳子,取出凝胶放入电泳槽内。
-
将电泳缓冲液加入电泳槽中,电泳缓冲液没过凝胶 为宜。
-
将扩增得到的 PCR 产物与凝胶载样缓冲液(内含指示剂)混合,再用微量移液器将混合液缓慢注入凝胶的加样孔内。留一个加样孔加入指示分子大小的标准参照物。
-
接通电源,根据电泳槽阳极至阴极之间的距离来设定电压,一般为 。待指示剂前沿迁移接近凝胶边缘时,停止电泳。
-
取出凝胶置于紫外灯下观察和照相。
注意
-
为避免外源 DNA 等因素的污染,PCR 实验中使用的微量离心管、枪头和蒸馏水等在使用前必须进行高压灭菌处理。
-
该实验所需材料可以直接从公司购买,缓冲液和酶应分装成小份,并在 储存。使用前,将所需试剂从冰箱拿出,放在冰块上缓慢融化。
-
在向微量离心管中添加反应组分时,每吸取一种试剂后,移液器上的枪头都必须更换。
蛋白质工程的原理和应用
人胰岛素在使用过程中容易聚合成多分子形式,阻断胰岛素从注射部位进入血液,从而延缓其降血糖的作用。另外,胰岛素进入血液循环后容易被降解,患者需要反复注射。上述两方面均与胰岛素分子的氨基酸序列有关。借助蛋白质工程可以定向设计、改变这些氨基酸,降低其聚合作用,使胰岛素快速发挥作用,同时可提高胰岛素在患者体内的稳定性。
重组 DNA 技术使得分离、克隆任何天然存在的基因并令其在特定受体细胞中表达成为可能,这项技术所使用的目的基因均是天然存在的,因而表达出的产物仍为天然蛋白质。天然蛋白质是生物在长期进化过程中形成的,它们的结构和功能符合特定物种适应特殊环境而生存的需求,但不一定都能完全符合人类生产和生活的需要。比如,人类常规主食(尤其是水稻和玉米)中的必需氨基酸(如赖氨酸)含量偏低;工业用酶在高温和有机溶剂生产环境中容易失活;药物蛋白多肽在机体内半衰期较短等。
以蛋白质结构与功能相统一的生命观念为指导,采用重组 DNA 技术在 DNA 分子水平上改变基因的序列和结构,可以对天然蛋白进行理性改造,甚至可以设计并制造出全新的非天然蛋白,以满足人类生产与生活的需求。这种由人为突变或设计基因进而操纵蛋白质结构和性质的过程,称为蛋白质工程,又称第二代基因工程。
蛋白质工程包含特定基因的定点突变和定向进化。定点突变是指改变 DNA 的特定序列;而定向进化是指随机改变 DNA 的序列,然后 选择具有特定功能的突变蛋白。基因工程涉及天然基因的重组,表达产物的结构和功能基本上没有改变;而蛋白质工程旨在对蛋白质的结构和功能进行设计和改造,获得性能更符合人类需求的蛋白质。蛋白质设计从预期的生物学功能出发,去改造蛋白质的结构,充分体现了结构和功能的统一。
蛋白质工程包括修饰改造天然蛋白和设计制造全新蛋白两个方面。修饰改造天然蛋白存在两大类操作策略,即基因的定点突变和定向进化。在 DNA 水平上改变蛋白质特定位点的氨基酸序列,称为基因的定点突变;而在 DNA 水平上随机改变蛋白质任一位点的氨基酸序列,则称为基因的定向进化。修饰改造天然蛋白一方面可实现氨基酸序列改造,优化其功能;另一方面还可以确定多肽链中某个氨基酸在蛋白质结构和功能上的作用,以收集有关氨基酸线性序列与其空间构象和生物功能之间的对应关系,为设计制作新型的突变蛋白提供理论依据。因此,修饰改造天然蛋白实质上是一个模拟自然界进化与适应的循环渐进过程,可将数千万年的自然进化历程缩短为数月、数周甚至数天的实验室过程。
我们知道,天然蛋白质合成的过程是按照中心法则进行的:
基因 $\to$ 表达(转录和翻译) $\to$ 形成具有特定氨基酸序列的多肽链 $\to$ 形成具有高级结构的蛋白质 $\to$ 行使生物功能
而蛋白质工程却与之相反,它的基本思路是:
从预期的蛋白质功能出发 $\to$ 设计预期的蛋白质结构 $\to$ 推测应有的氨基酸序列 $\to$ 找到并改变相对应的脱氧核苷酸序列(基因)或合成新的基因 $\to$ 获得所需要的蛋白质
借助蛋白质工程设计和改造新型突变蛋白,具有不可估量的经济和社会效益。让我们结合具体案例了解如何借助定点突变策略改造蛋白质的基因结构,进而产生突变蛋白的大致过程。
-
定点突变:研究表明,人胰岛素样生长因子的结构和性质与人胰岛素高度相似,但它不像人胰岛素那样容易聚合形成多分子。比较这两种蛋白质的结构发现,人胰岛素样生长因子在与人胰岛素 B 链对应的第 28 位和第 29 位氨基酸分别是赖氨酸和脯氨酸,而人胰岛素在这两处的氨基酸序列正好颠倒,因而推测这是人胰岛素形成多分子形式的关键原因。为了证实这一猜想,需要更换人胰岛素这两个位点的氨基酸顺序。
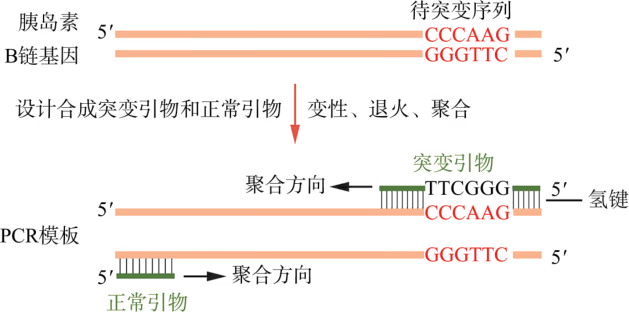
已知,人胰岛素 B 链基因中编码第 28 位脯氨酸和第 29 位赖氨酸的序列分别为 CCC 和 AAG。因此,只需在人胰岛素基因中将原来的"CCCAAG"序列改造成"AAGCCC",理论上便能在受体细胞中产生两处氨基酸序列颠倒的突变型人胰岛素。由于人胰岛素 B 链基因较短,按照设计好的编码序列进行化学合成,是获取这种突变型目的基因的首选方法。另外,也可采取图 3-17 所示的 PCR 方案获取突变型目的基因(PCR 方案对较长的目的基因编码序列更经济):首先人工化学合成含有突变序列的突变引物和不含突变序列的正常引物,然后使用这两种引物以人胰岛素基因为模板进行 PCR。研究证实,由这种突变的目的基因表达的突变型人胰岛素不再聚合成多分子形式。将这种突变型人胰岛素注射进入患者体内,降血糖速率大为提高,因而这种突变型人胰岛素被称为"单体速效胰岛素"(又称"赖脯胰岛素")。
L-天冬酰胺酶是治疗儿童白血病的有效药物,但在临床应用中因其并非人体蛋白常常引起过敏反应,因此降低免疫原性是该药研究开发的重要内容。采用定点突变技术将细菌来源的 L-天冬酰胺酶某位赖氨酸更换为丙氨酸,形成新型的 L-天冬酰胺酶突变蛋白,其免疫原性比天然酶蛋白下降 倍,且活性保持不变。
再如,心脑血管疾病引发的血栓(即血液中主要由蛋白质构成的固体颗粒)并发症严重威胁人类生命,全世界每年死于心脑血管疾病的人数高达 万人,居各种疾病死亡率首位。因此,研制高效、特异、安全的溶栓药物一直是药物开发领域的热门课题。临床上常将人组织型纤溶酶原激活剂(t-PA)用作溶解血栓、抢救心肌梗死(心脏血管被血栓堵塞)患者的特效药。然而,t-PA 存在体内半衰期短和诱发颅内出血等缺陷。为了提高 t-PA 临床应用的可行性,科研人员采用蛋白质工程的定点突变策略,开发了一系列体内半衰期长、血栓溶解效率高、颅内出血倾向大幅度降低的优质 t-PA 突变蛋白,如 TNK 型 t-PA。
-
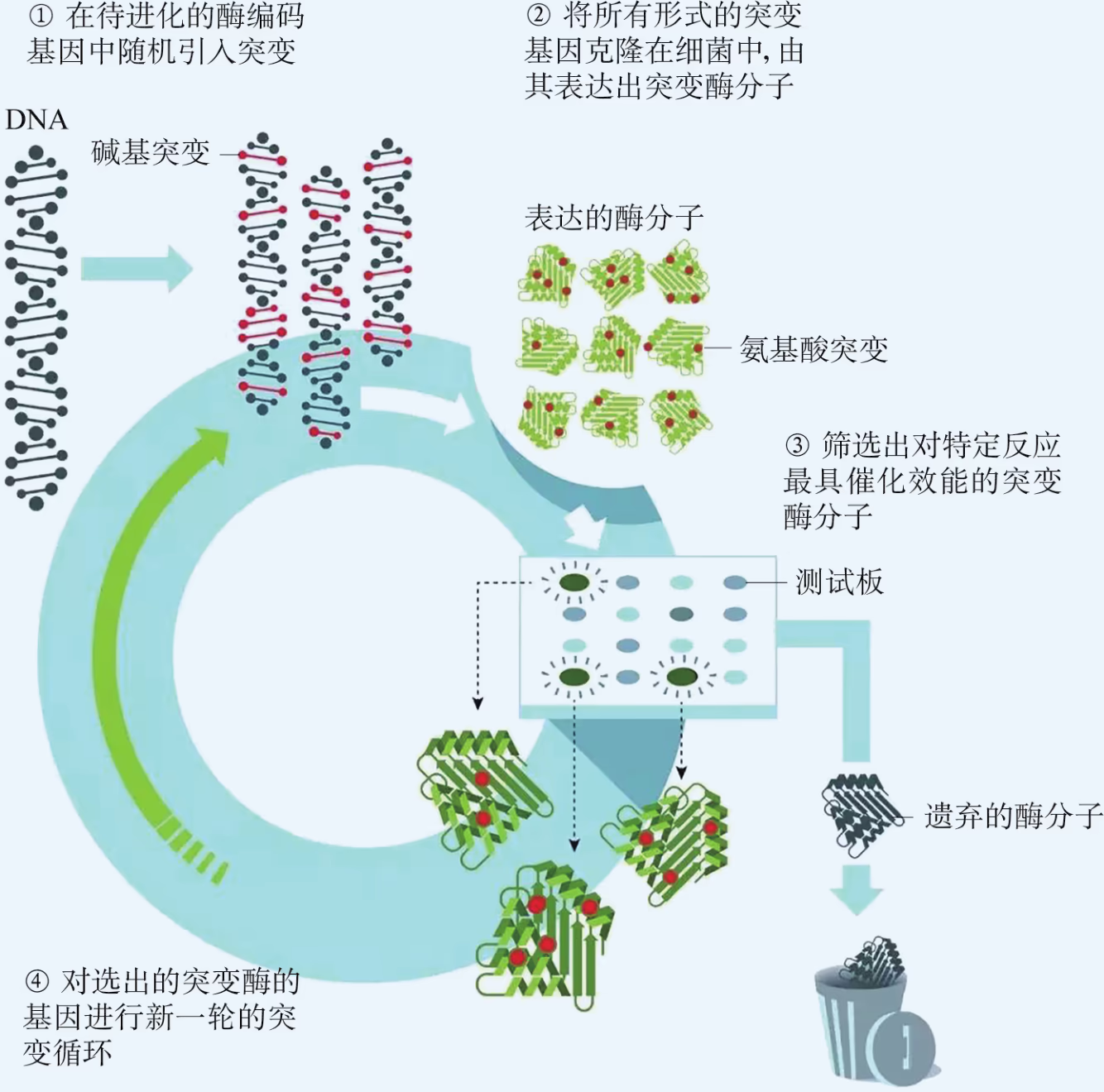
随机突变:随机突变以获得高效能酶分子。
-
定向进化:进化到今天的生物体基因都是最优的吗?我们能不能在实验室里模拟并加速自然进化过程呢?基因的定向进化有助于解答这些问题。这项技术的基本操作是在体外对特定基因实施随机突变,然后借助适当的筛选程序准确、迅速地获得所需要的突变基因。比如,针对某个特定的基因采用 PCR 技术进行扩增,在过程中通过改变反应条件(如使用低保真度的 DNA 聚合酶)故意让 DNA 复制发生随机错误,便能创建一系列随机突变的新基因序列;然后再将这些新基因序列导入合适的受体细胞中,表达出相应的一系列随机突变蛋白;最后,根据我们的需求从中筛选出具有特定优良功能或结构的目的蛋白。
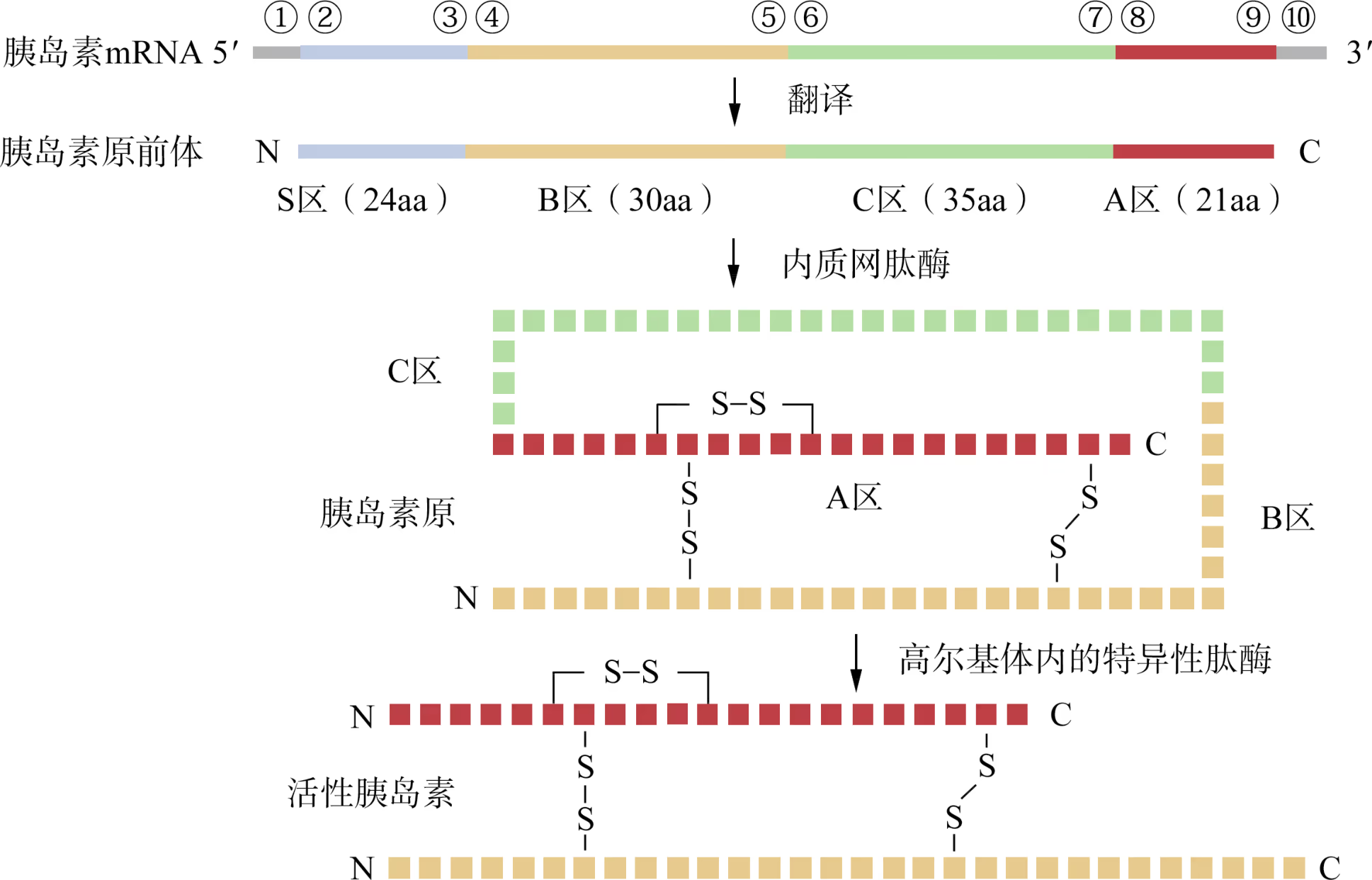
在人类胰岛 β 细胞中,胰岛素基因最初表达的是由 110 个氨基酸残基(aa)构成的胰岛素原前体分子,其 N 端的 S 区在新生多肽链进入内质网腔后被切除,形成无活性的胰岛素原(即 B-C-A),其中 A 区与 B 区借助二硫键(共价键)相连。随后,胰岛素原被转运至高尔基体。当机体接到胰岛素的需求指令后,高尔基体内的肽酶再切除胰岛素原中的 C 区,形成 A 区与 B 区相连的活性胰岛素(三个二硫键的正确搭配为胰岛素活性所必需)。
鉴于活性胰岛素仅含 A 区和 B 区,科研人员最初设计的人胰岛素基因工程生产技术路线是:首先化学合成 A 区和 B 区编码序列,然后分别与相同的质粒连接,构建成表达载体 p Ⅰ A1 和 p Ⅰ B1。将这两种重组 DNA 分子分别导入大肠杆菌,便能分别表达人胰岛素的 A 区和 B 区。经分离纯化后,将 A 区与 B 区混合,使其自然形成三个二硫键。由此技术路线(即 AB 表达法)生产的重组人胰岛素成本高昂。
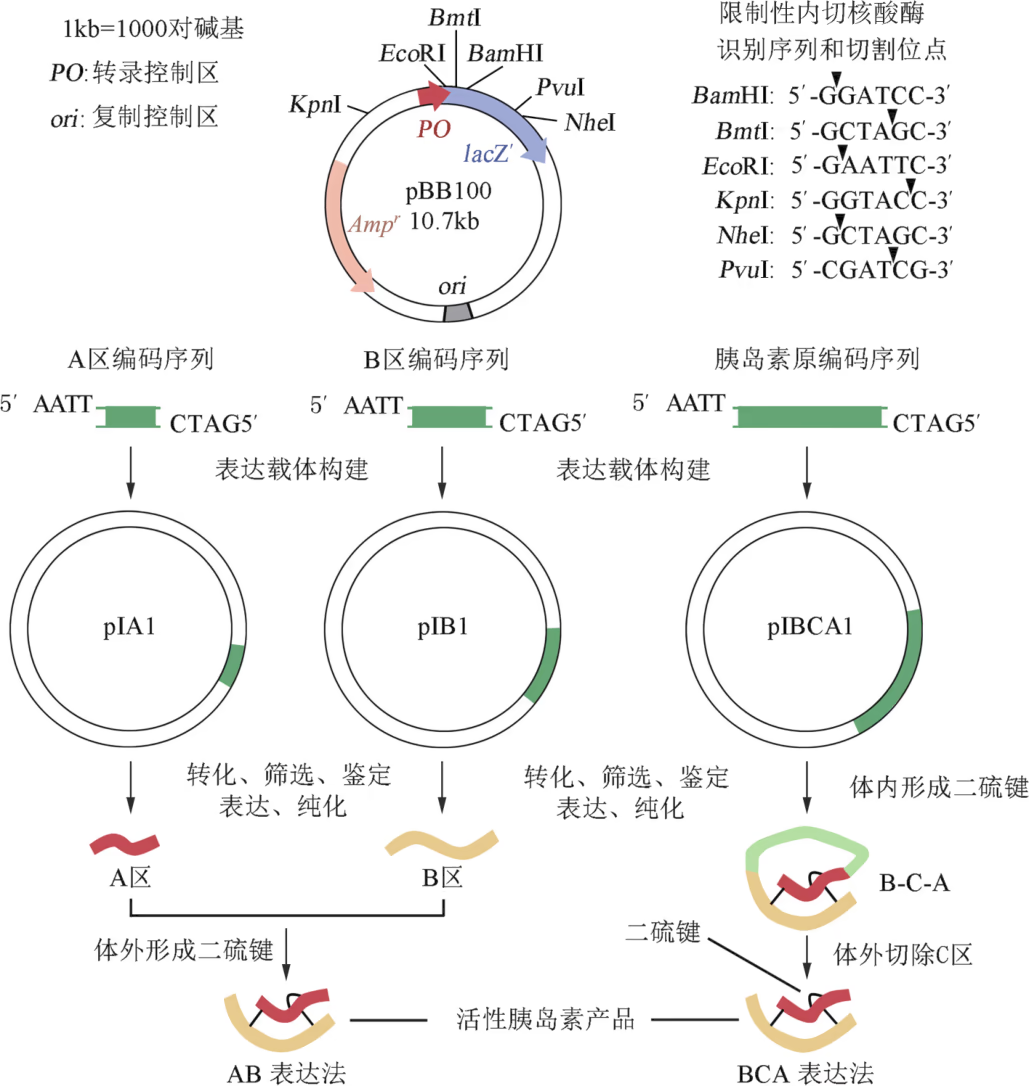
为了降低生产成本,科研人员重新审视了 C 区的作用,他们认为在胰岛 β 细胞内 C 区很可能起到正确组装 A 区和 B 区的"脚手架"作用。为了验证这一假说,他们以胰岛 β 细胞的 mRNA 为原料,采用 PCR 技术获取胰岛素原编码序列,然后依照上述工艺制备 B-C-A,最后再用特殊的酶模拟细胞内的过程将 C 区切除。结果表明,由此技术路线(即 BCA 表达法)生产的人胰岛素基因工程产品成本大幅度降低。
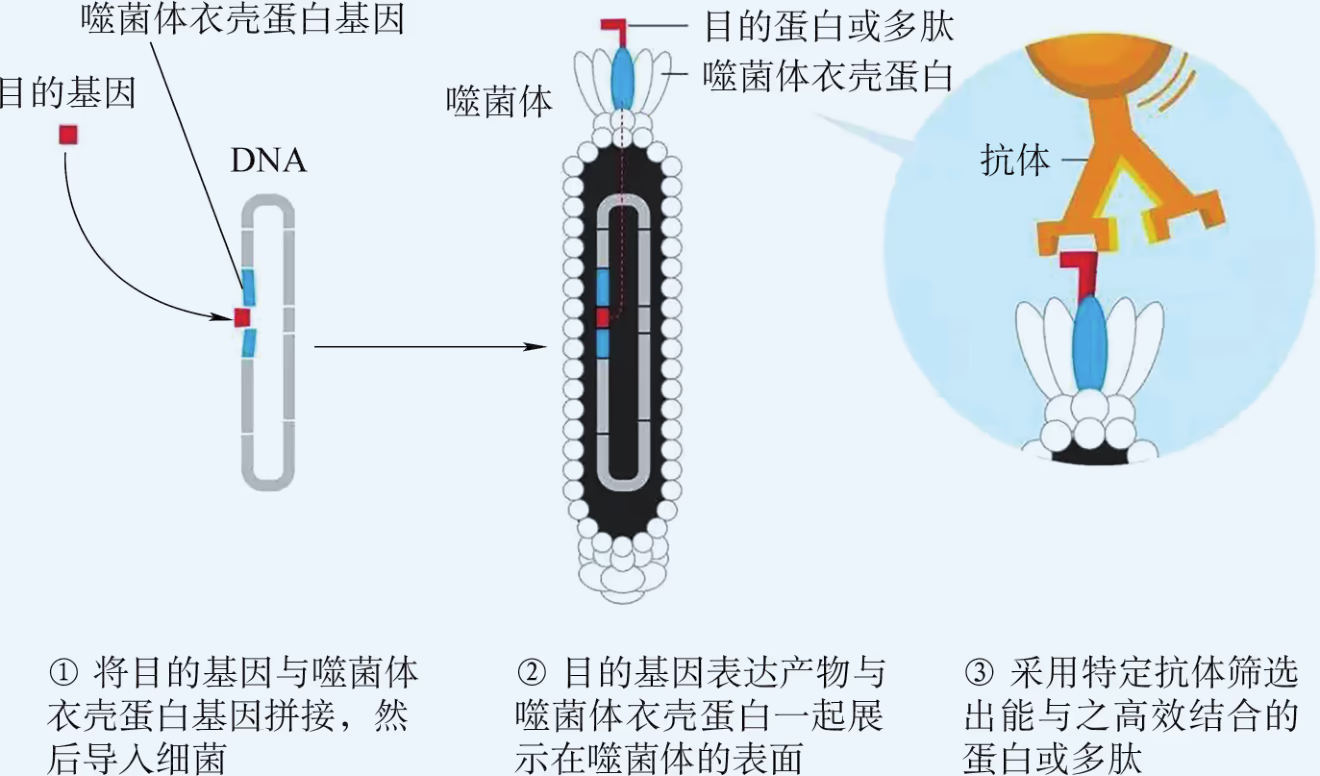
设计制造全新蛋白需要根据已经掌握的蛋白质结构与功能的关系,借助特殊的电脑程序自行设计、拼装感兴趣的全新基因序列,然后借助重组 DNA 技术使之表达,进而检测这种全新蛋白的性能是否能达到设计标准。
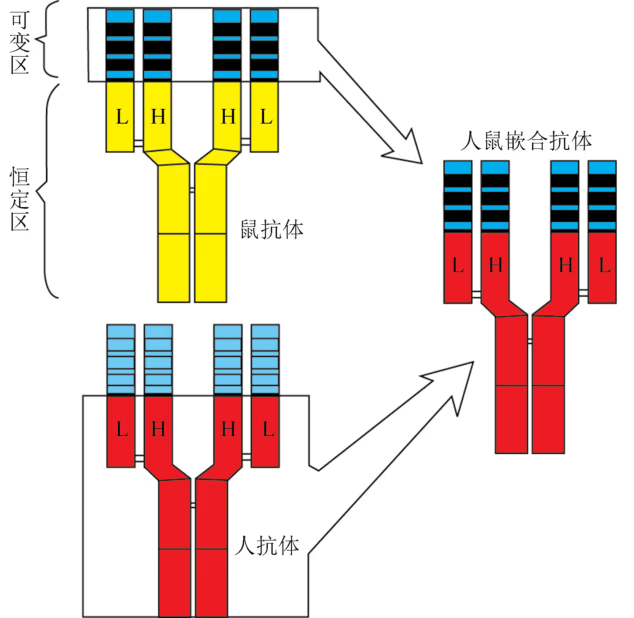
小鼠单克隆抗体的制备比较简单,但这种鼠源性的单克隆抗体会被人的免疫系统排斥,不能直接用于人体。嵌合抗体(chimeric antibody)分子的可变区是由鼠抗体的可变区基因所编码,恒定区则由人抗体的恒定区基因编码,这种嵌合抗体的抗原性显著下降,而抗体的特异识别功能没有丧失。目前,已有多种嵌合抗体用于临床治疗。
蛋白质工程经常要借助计算机来建立蛋白质的三维结构模型;要制备蛋白质晶体,然后通过 X 射线衍射技术,分析晶体的结构;要用到基因的定点突变技术来进行碱基的替换。蛋白质工程是一项难度很大的工程,主要是因为蛋白质发挥功能必须依赖于正确的高级结构,而这种高级结构往往十分复杂(图 3-18)。科学家要设计出更加符合人类需要的蛋白质,还需要不断地攻坚克难。我们相信,随着科学技术的深入发展,蛋白质工程将会给人类带来更多的福祉。

蛋白质是生命活动的主要承担者,蛋白质结构改变会直接影响其功能活性。蛋白质工程技术已经能够对蛋白质稳定性、最适 pH 范围、酶的活性以及底物特异性等进行预期的设计和改造。提高胰蛋白酶的活性与稳定性胰蛋白酶在医药、轻工业、科学研究等方面有广泛的应用价值,但天然胰蛋白酶活性低,稳定性差。科学家通过选择胰蛋白酶第 216 位和第 226 位的甘氨酸进行定点突变,获得了催化活性更高的胰蛋白酶;通过对第 117 位的精氨酸进行定点突变,获得了稳定性更高的胰蛋白酶。并以此为手段,进一步研究了胰蛋白酶底物专一性和胰蛋白酶结构与功能的关系。
改变金属硫蛋白对重金属的亲和力金属硫蛋白(MT)中丰富的巯基对金属离子有很高的亲和性,对于抵抗重金属中毒及维持某些必需元素在体内的代谢平衡具有重要意义。MT 一般由 α 和 β 两个功能区域组成。科学家将 α 和 β 两个功能区域拆分后,用 α 功 能区域代替 β 功能区域,构建出含两个 α 功能区域的 MT 突变体 α-α,然后将 MT 突变体 α-α 转入烟草体内,使烟草具有较高的重金属抗性。
延长白细胞介素-2 的作用时间:白细胞介素-2(IL-2)是一种细胞因子,能够促进 T 淋巴细胞增殖以及增强某些免疫细胞的活性,已被广泛运用于各种肿瘤和病毒感染疾病的治疗。IL-2 作为临床药物在血浆中的作用时间较短,治疗期间需要频繁给药。科学家通过蛋白质工程,将长效人血清白蛋白(HSA)与 IL-2 重组,获得了一种新的融合蛋白(HSA/IL-2),这种融合蛋白既具有普通 IL-2 的生物活性,作用时间又有所延长,可明显提高 IL-2 的疗效。
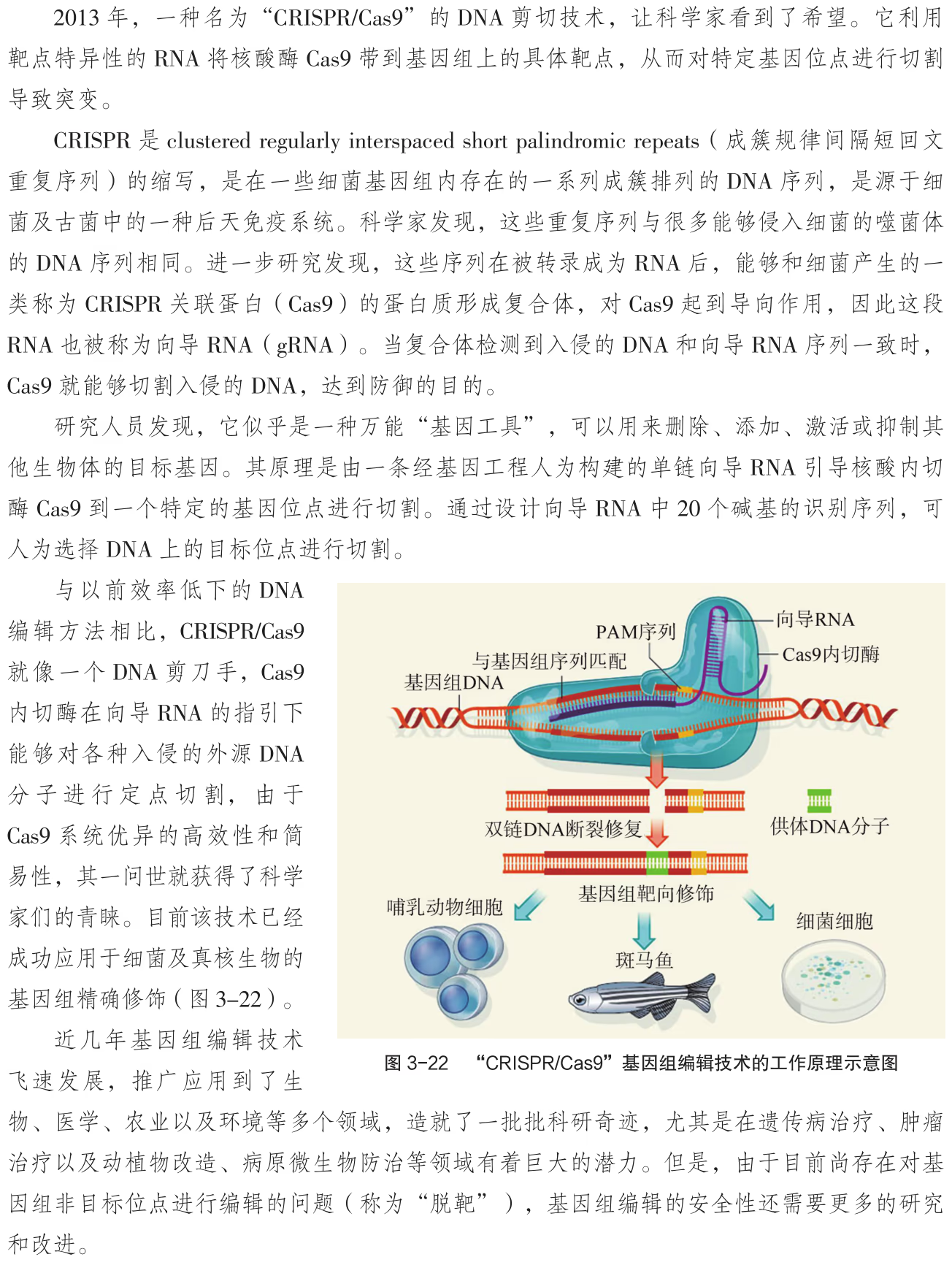
"CRISPR/Cas9"基因组编辑技术
相对小分子药物而言,蛋白质药物具有活性高、特异性强、毒性低、生物功能明确、有利于临床应用等特点。目前上市的蛋白质工程药物常见种类有:基因工程抗体药物,多肽类激素药(包括人胰岛素、人生长激素、卵泡刺激激素等),人造血因子(包括重组人促红细胞生成素、粒细胞 / 单核细胞集落刺激因子等),人细胞因子(包括α干扰素、β干扰素等),人血浆蛋白因子(包括重组人凝血因子、t-PA 等),人骨形成蛋白,融合蛋白,外源重组蛋白等。
蛋白质工程汇集了当代分子生物学等学科的一些前沿领域的最新成就,将蛋白质的研究推进到崭新的时代,为蛋白质在工业、农业和医药方面的应用开拓了诱人的前景。蛋白质工程开创了按照人类意愿改造、创造符合人类需要的蛋白质的新时期。
基因工程的基本操作程序
基因工程是一种重组 DNA 技术,即按照人们预先设计的蓝图,通过基因操作,将目的基因或 DNA 片段与合适的载体结合,然后转入受体细胞,通过复制、转录、翻译,使转基因生物获得新的遗传性状。基因工程可以打破物种之间的生殖隔离,使基因在不同生物之间进行交流,从而赋予生物新的遗传特性。
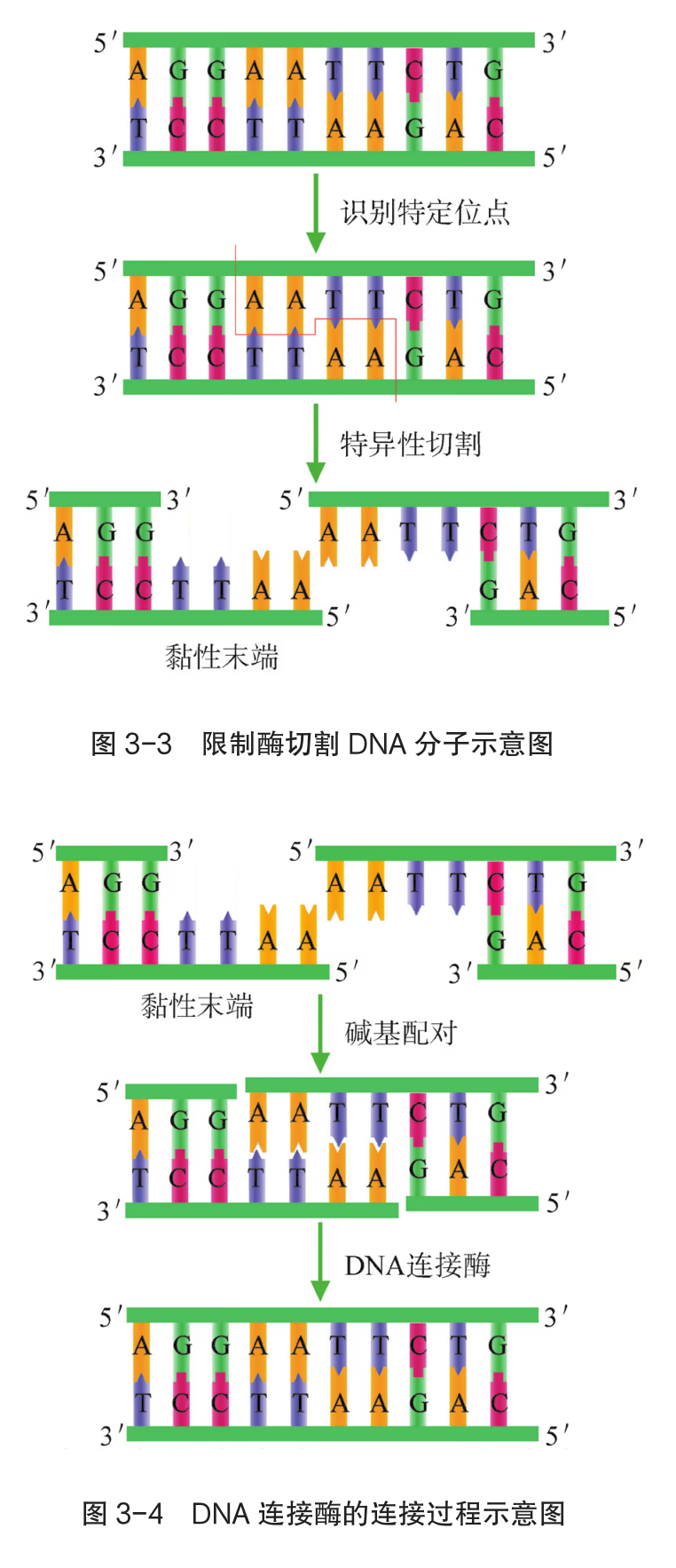
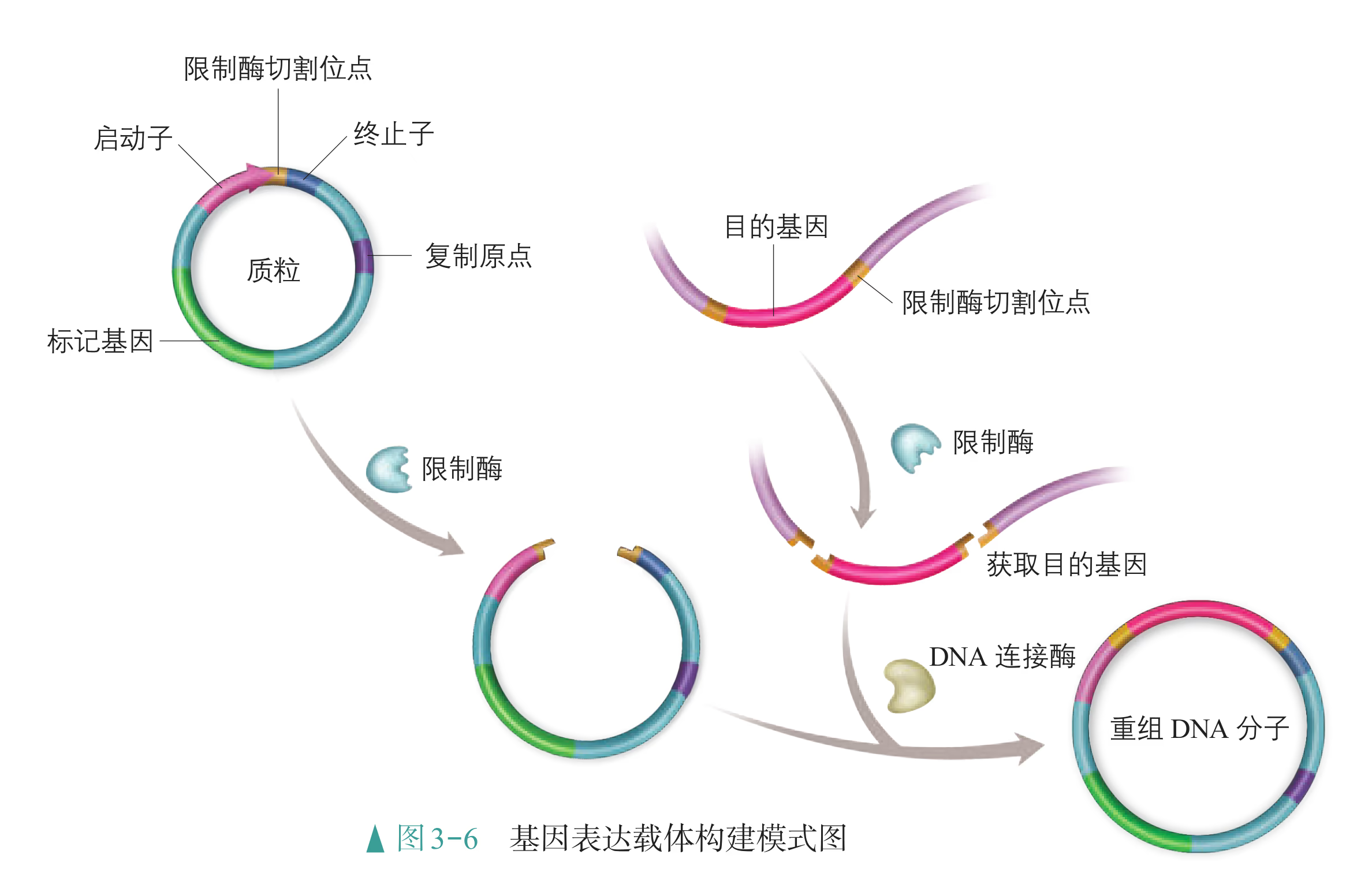
重组 DNA 过程中需要三种基本的工具,即切割 DNA 分子的"基因剪刀"、将 DNA 分子片段连接起来的"基因针线"、将体外基因导入受体细胞的"运载工具"。 "基因剪刀" ------限制性内切核酸酶:限制性内切核酸酶(restriction endonuclease),又称限制酶(restriction enzyme)。迄今为止,科学家已从微生物体内分离出了几千种不同的限制酶,每一种限制酶能够识别 DNA 分子中特定的脱氧核苷酸序列,并在特定位置将 DNA 分子"切割"开。大部分 DNA 分子被切割后,切口两端会分别留下一段单链末端,叫作黏性末端(图 3-3)。有些限制酶切开 DNA 时,双链末端平齐而无突出单链,叫作平端。 "基因针线" ------DNA 连接酶:将切割下来的 DNA 片段拼接成新的 DNA 分子,要靠 DNA 连接酶(DNA ligase)来实现。当两个 DNA 片段的黏性末端彼此靠拢,相互识别,它们的碱基完成互补配对后,DNA 连接酶就把它们之间的缝隙"缝合"起来(图 3-4)。有些连接酶可以连接 DNA 片段的平端。
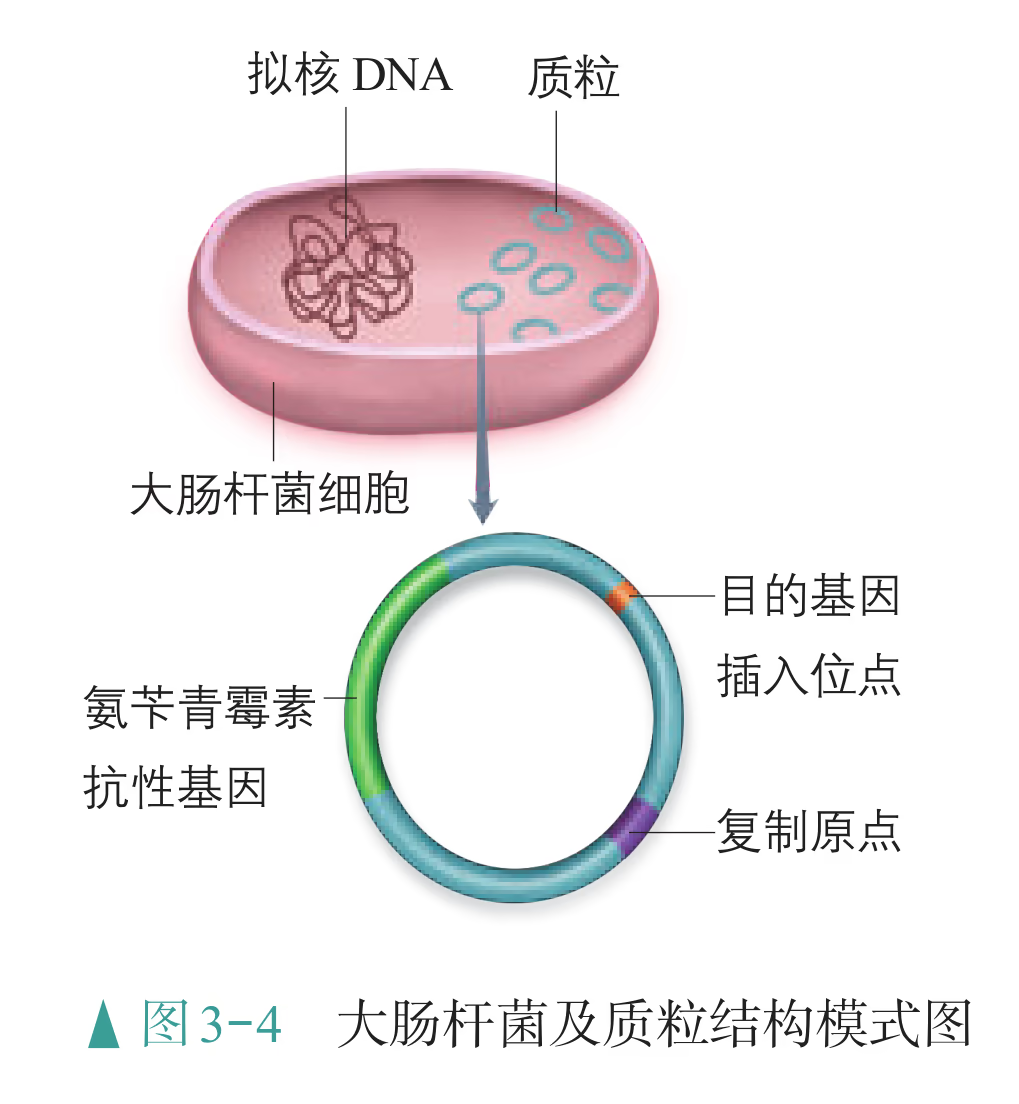
- "运载工具" ------载体:将外源基因送入受体细胞,需要运载工具,这种运载工具称为载体(vector)。常用的载体有质粒、噬菌体、动植物病毒等。其中,质粒是一种较为理想的载体,它是一种很小的环状 DNA 分子,具有自我复制能力。质粒 DNA 分子上有一个至多个限制酶切割位点,并含有一些抗生素抗性基因(图 3-5),如四环素抗性基因、氨苄青霉素抗性基因等,这些基因可以作为鉴定和筛选重组 DNA 的标记基因
::: {.grid}
 :::
:::
一、DNA 重组需要三种基本工具
通过模拟 DNA 重组操作,你可以发现若将人胰岛素基因插入环形 DNA 分子的特定位点处,必须进行的操作步骤是"切"和"接"。实现这一精确的操作过程至少需要三种分子工具,即准确切割 DNA 分子的分子手术刀、将 DNA 片段再连接起来的分子缝合针和将体外重组好的 DNA 分子导入受体细胞的分子运输车。
-
限制性内切核酸酶:切割 DNA 分子的工具是限制性内切核酸酶,简称限制酶。这类酶主要是从原核生物中分离纯化出来的。迄今分离的限制酶有数千种,许多已经被商业化生产。它们能够识别双链 DNA 分子的特定核苷酸序列,并且使每一条链中特定部位的磷酸二酯键断开。
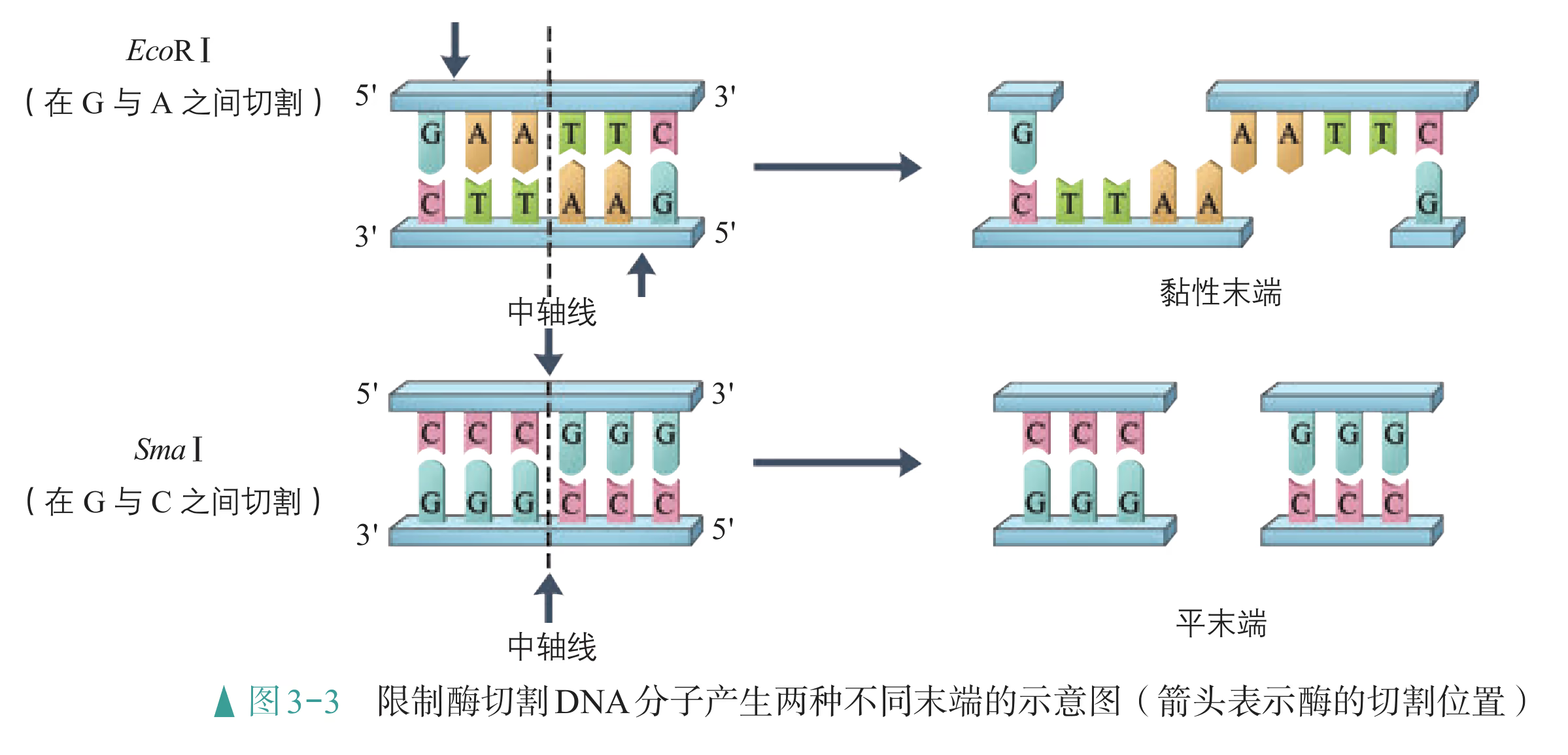
大多数限制酶的识别序列由 个核苷酸组成。例如,EcoRⅠ、SmaⅠ限制酶的识别序列均为 个核苷酸,也有少数限制酶的识别序列由 个、 个或其他数量的核苷酸组成。DNA 分子经限制酶切割产生的 DNA 片段末端通常有 两种形式(黏性末端和平末端)。当限制酶在它识别序列的中心轴线(图中虚线)两侧将 DNA 分子的两条链分别切开时,产生的是黏性末端;当限制酶在它识别序列的中心轴线处切开时,产生的是平末端。
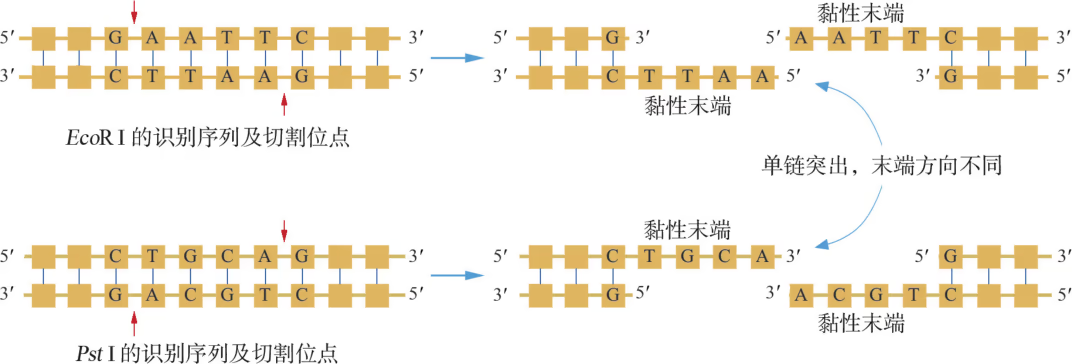
20 世纪 60~70 年代,限制性内切核酸酶的发现使人们能够对双链 DNA 分子进行精确切割。目前,在细菌中已发现了上千种限制性内切核酸酶,每一种限制性内切核酸酶识别双链 DNA 特定的序列,长度通常为 4~8 个碱基对(bp),称为识别序列。例如,限制性内切核酸酶 EcoR Ⅰ 的识别序列为 5'-GAATTC-3';而 Pst Ⅰ 的识别序列则为 5'-CTGCAG-3'。如图 3-7 所示,当限制性内切核酸酶与双链 DNA 的识别序列结合后,便能断裂识别序列内部或两侧相邻两个脱氧核苷酸之间(即切割位点)的化学键,从而切开 DNA 分子的两条链。无论 EcoR Ⅰ 还是 Pst Ⅰ ,切割后形成的双链 DNA 片段末端会呈单链突出,但两种酶切产物的单链突出方向有所不同。从图中还可以看到,如果线形 DNA 分子两个端点的单链突出部分呈序列互补性且方向相同,那么它们便能在较低温度下重新"粘"在一起(即互补碱基之间形成氢键),这样的单链末端称为黏性末端。
在微生物培养中,人们观察到生长在大肠杆菌某些菌株(如 K 株)中的噬菌体不能在其他菌株(如 B 株)中存活,或者生长能力受到严重限制。于是便产生这样的疑问:大肠杆菌不同菌株对噬菌体生长的这种限制是由什么决定的?为了解答这一问题,研究人员作了如下的实验设计:将 B 株或 K 株分别与其他大肠杆菌菌株按一定比例混合培养,然后再用噬菌体感染这种细菌混合培养物。结果显示,从其他大肠杆菌菌株中释放出来的噬菌体感染 B 株或 K 株的能力也大打折扣。进一步深入分析发现,这种限制效应是由细菌株产生的一种酶所引起的,这种酶能识别"外源"噬菌体 DNA 中的特异性位点并裂解之,因而就被命名为"限制性内切核酸酶"。更为奇妙的是,为了保护其自身的 DNA 免遭自己产生的限制性内切核酸酶切割,细菌还会表达一种 DNA 甲基化酶,催化限制性内切核酸酶识别序列内某种碱基的甲基化(即修饰效应),从而使其抗限制性内切核酸酶的切割。不难想象,如果外源入侵的 DNA 在此类细菌菌株中偶然站住脚,同样会被宿主细胞内的甲基化酶所修饰,从而形成对限制性内切核酸酶切割作用的"免疫"。细菌的这种限制 - 修饰机制相当于脊椎动物体内的先天性免疫系统,不具备免疫记忆功能。
-
DNA 连接酶:将切下来的 DNA 片段拼接成新的 DNA 分子,是靠 DNA 连接酶来完成的。1967 年,世界上几个实验室几乎同时发现了一种能够将两个 DNA 片段连接起来的酶,称之为 DNA 连接酶(DNA ligase)。在基因工程操作中,DNA 连接酶主要有两类:一类是从大肠杆菌中分离得到的,称为 E.coli DNA 连接酶;另一类是从 T4 噬菌体中分离出来的,称为 T4 DNA 连接酶。这两类酶都能将双链 DNA 片段"缝合"起来,恢复被限制酶切开的磷酸二酯键,但 E.coli DNA 连接酶连接具有平末端的 DNA 片段的效率要远远低于 T4 DNA 连接酶。
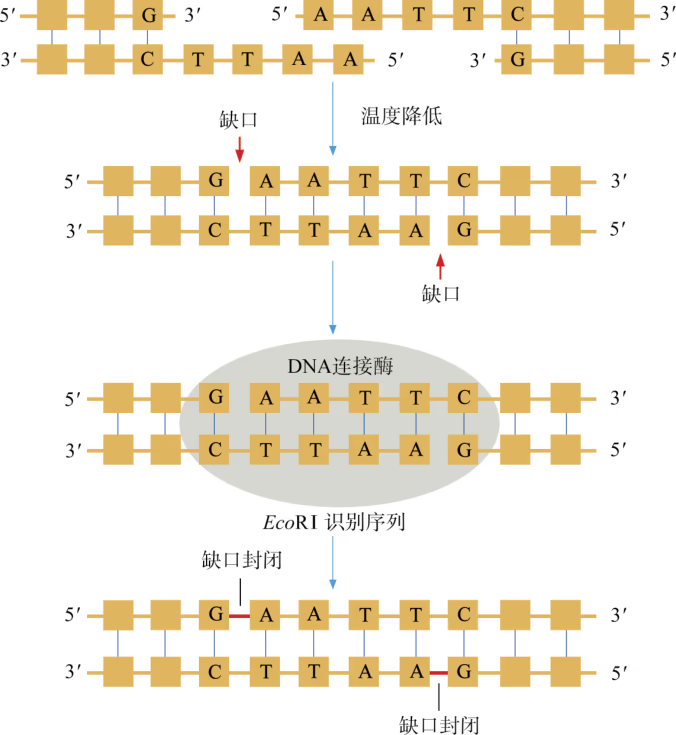
DNA 连接酶 黏性末端之间在低温下形成的氢键并不牢固,若要使两个黏性末端牢固地连为一体,便需要使用另一种工具:DNA 连接酶。大多数生物的 DNA 连接酶能封闭双链 DNA 分子中的单链"缺口",即一条链上相邻两个脱氧核苷酸之间断开的化学键(共价键),因此特别适合将两个黏性末端牢固地连为一体。
-
将外源基因导入受体细胞中,为什么要使用载体呢?大部分 DNA 片段(尤其是目的基因)并不具备可遗传的自我复制能力,因为缺少复制所必需的特定 DNA 序列。而且,即便一个 DNA 片段能在原供体细胞中复制,这种复制能力一般情况下也难以在受体细胞中正常发挥。因此,要让外源 DNA 在受体细胞中复制,就需要一种合适的运载工具为其提供在受体细胞中复制的能力,这种用于重组 DNA 技术的运载工具称为载体。也就是说,任何具备在受体细胞中独立复制性能的 DNA 分子,理论上均可用作基因工程的载体。
怎样才能将外源基因送入细胞呢?通常是利用质粒(plasmid)作为载体(vector),将基因送入细胞。质粒是一种裸露的、结构简单、独立于真核细胞细胞核或原核细胞拟核 DNA 之外,并具有自我复制能力的环状双链 DNA 分子。质粒 DNA 分子上有一个至多个限制酶切割位点,供外源 DNA 片段(基因)插入其中。携带外源 DNA 片段的质粒进入受体细胞后,能在细胞中进行自我复制,或整合到受体 DNA 上,随受体 DNA 同步复制。在基因工程操作中,真正被用作载体的质粒,都是在天然质粒的基础上进行过人工改造的。这些质粒上常有特殊的标记基因,如四环素抗性基因、氨苄青霉素抗性基因等,便于重组 DNA 分子的筛选(图 3-4)。
生物界至少存在两大类能独立于拟核 DNA 或染色体 DNA而自主复制的小型 DNA 分子,即广泛存在于微生物细胞内的质粒 DNA 以及存在于大多数生物体内的病毒 DNA。大多数质粒是一类双链环状 DNA 分子(图 3-9)。质粒并非其宿主正常生长所必需,但大都携带抗生素抗性等基因,因而能帮助宿主抵御环境不利因素的影响。除了质粒 DNA 和病毒 DNA 外,生物体内还存在其他能独立于染色体 DNA 而自主复制的小型 DNA,它们的结构与功能均有别于质粒 DNA和病毒 DNA。病毒不仅能在宿主细胞中自主复制,还能通过感染将其 DNA 高效导入宿主细胞中。
在基因工程中使用的载体除质粒外,还有噬菌体、动植物病毒等。它们的来源不同,在大小、结构、复制方式以及可以插入外源 DNA 片段的大小上也有很大差别。这些基因工程载体,相当于一种运输工具,因此将它们比喻为"分子运输车"。
DNA、RNA、蛋白质和脂质等在物理和化学性质方面存在差异,可以利用这些差异,选用适当的物理或化学方法对它们进行提取。例如,DNA 不溶于酒精,但某些蛋白质溶于酒精,利用这一原理,可以初步分离 DNA 与蛋白质。DNA 在不同浓度的 溶液中溶解度不同,它能溶于 的 溶液。在一定温度下,DNA 遇二苯胺试剂会呈现蓝色,因此二苯胺试剂可以作为鉴定 DNA 的试剂。
二、PCR 是获取目的基因的主要方法
目的基因是人们为了达到基因工程特定目标而导入受体细胞的基因。例如,在规模化生产人胰岛素的基因工程中,人胰岛素基因就是目的基因。从供体生物的 DNA 中有效分离和克隆目的基因是基因工程各项产业化应用的前提,也是基因工程项目实施的首要阶段。
在基因工程的设计和操作中,用于改变受体细胞性状或获得预期表达产物等的基因就是目的基因。根据不同的需要,目的基因是不同的,它主要是指编码蛋白质的基因,如与生物抗逆性、生产药物、毒物降解、工业用酶等相关的基因。随着测序技术的发展,以及序列数据库(如 GenBank)、序列比对工具(如 BLAST)等的应用,越来越多的基因的结构和功能为人们所知,这也为科学家找到合适的目的基因提供了更多的机会和可能。
采用何种方法分离获得特定的目的基因,取决于我们对目的基因背景知识的了解程度。如果目的基因的部分序列已知,可采用聚合酶链式反应(Polymerase Chain Reaction,PCR);如果目的基因的完整序列已知,还可采用化学合成法;如果基因序列未知,则采用基于大规模筛选的方法获得。PCR 是聚合酶链式反应的缩写。它是一项根据 DNA 半保留复制的原理,在体外提供参与 DNA 复制的各种组分与反应条件,对目的基因的核苷酸序列进行大量复制的技术(表 3-1)。
PCR 是一项模拟细胞内 DNA 复制过程的 DNA 体外扩增技术,由美国科学家穆勒斯(K. B. Mullis)于 20 世纪 80 年代中期首创。利用这项技术可由微量的 DNA 样品特异、高效、准确地扩增特定区域的 DNA 序列。采用 PCR 扩增技术获取目的基因需要 DNA 模板、引物、DNA 聚合酶、四种脱氧核苷三磷酸(dNTP)以及缓冲液系统。由于几乎所有生物来源的 DNA 聚合酶只能在一段与模板 DNA 发生碱基互补配对的单链核酸(称为引物)存在的条件下,才能催化 DNA 链的延伸聚合反应,因此, PCR 扩增目的基因的前提条件是目的基因两侧的序列已知,以便人工化学合成 DNA 引物。PCR 的基本程序如图 3-10 所示。
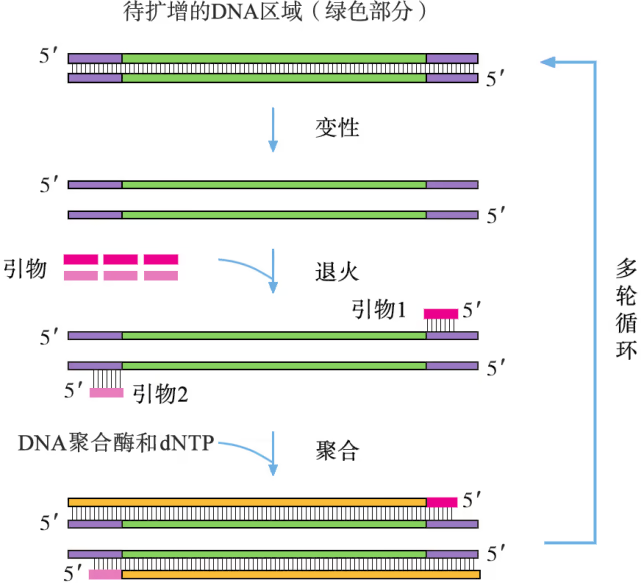
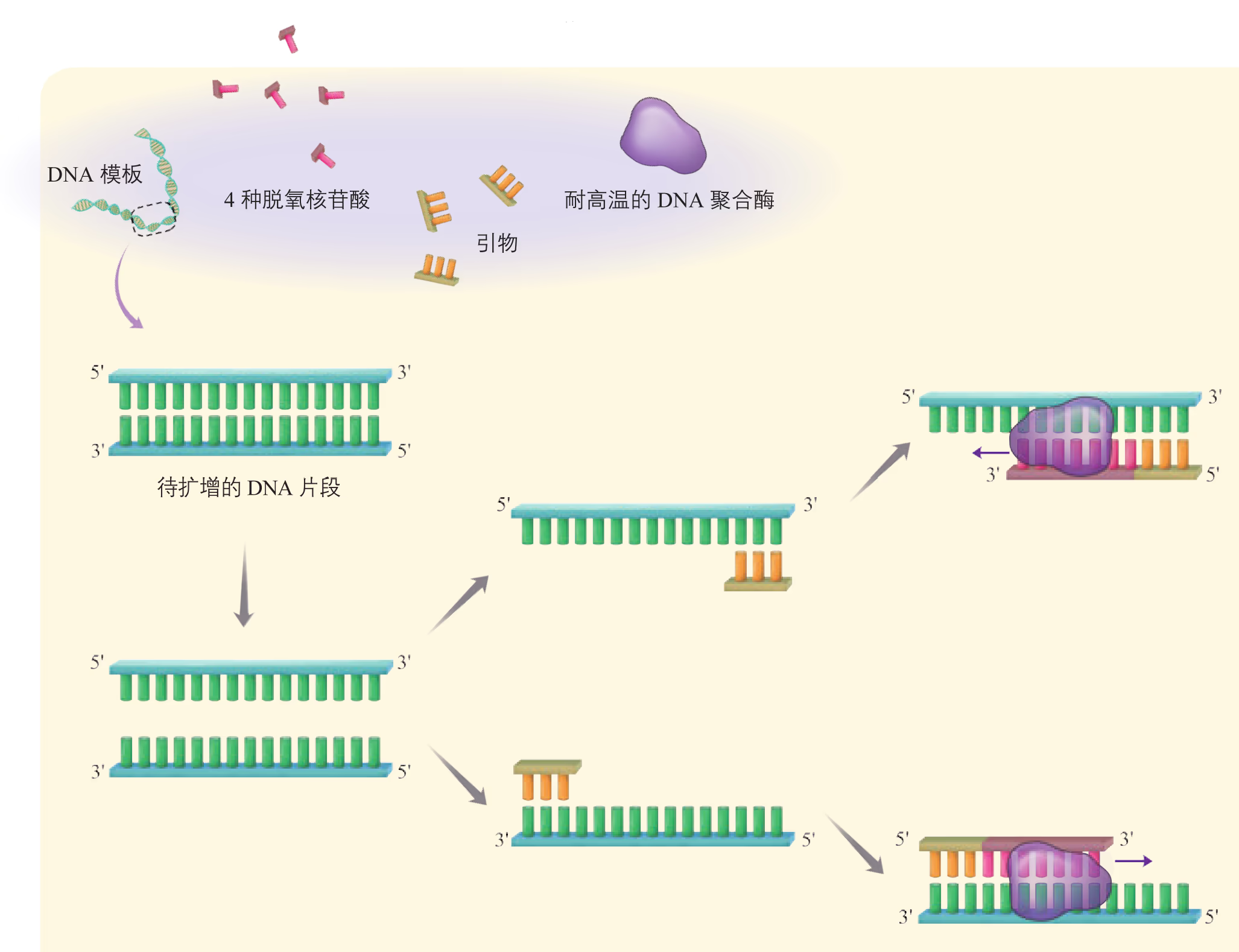
PCR 反应需要在一定的缓冲液中才能进行,需提供 DNA 模板,分别与两条模板链结合的 2 种引物,4 种脱氧核苷酸和耐高温的 DNA 聚合酶;同时通过控制温度使 DNA 复制在体外反复进行。扩增的过程是:目的基因 DNA 受热变性后解为单链,引物与单链相应互补序列结合;然后以单链 DNA 为模板,在 DNA 聚合酶作用下进行延伸,即将 4 种脱氧核苷酸加到引物的 端,如此重复循环多次。由于延伸后得到的产物又可以作为下一个循环的模板,因而每一次循环后目的基因的量可以增加一倍,即成指数形式扩增(约为 ,其中 为扩增循环的次数)。每次循环一般可以分为变性、复性和延伸三步(图 3-5)。
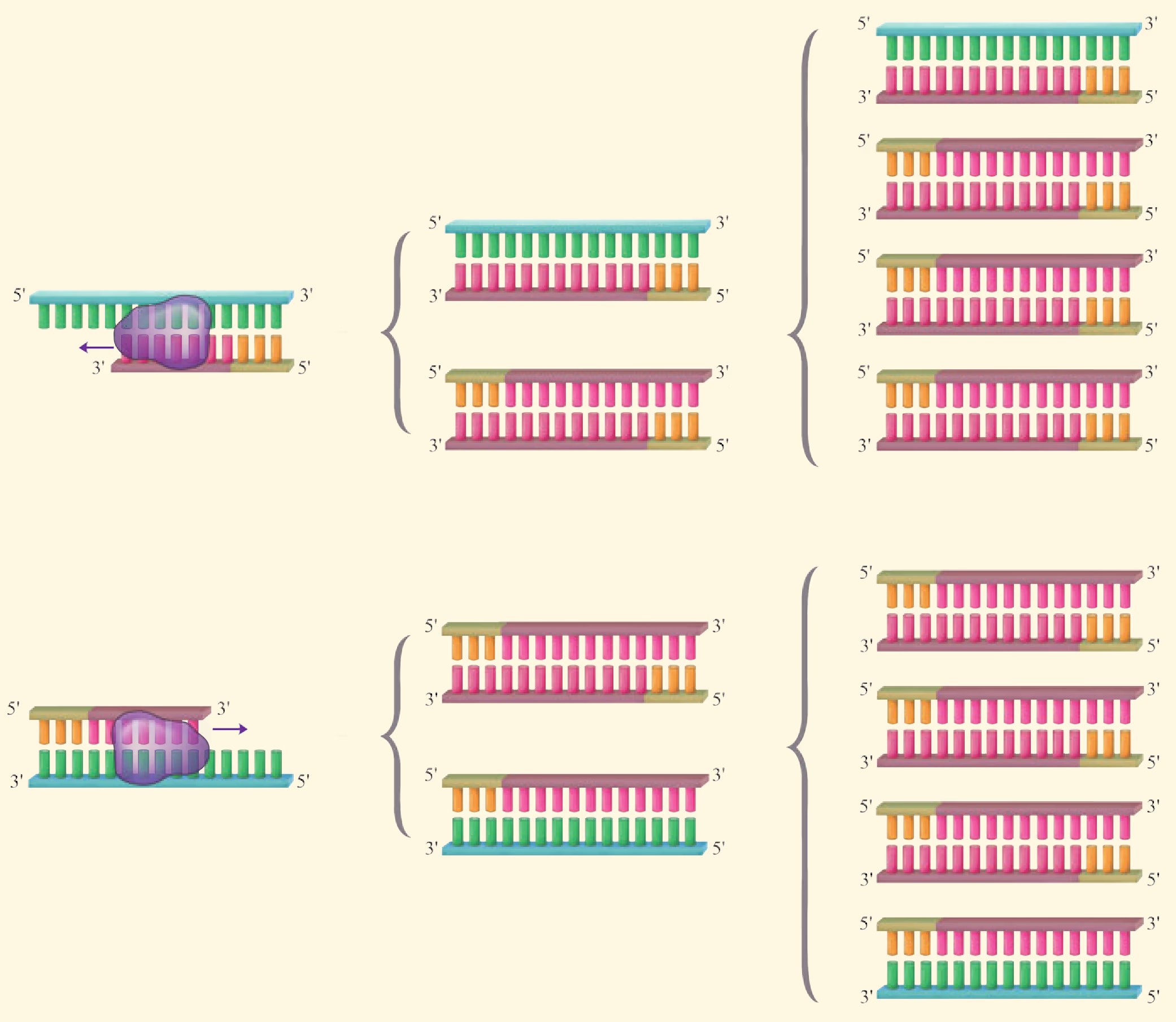
-
变性:当温度超过 时,双链 DNA 解聚为单链。
-
复性(退火):当温度下降到 左右时,两种引物通过碱基互补配对与两条单链 DNA 结合。
-
延伸(聚合):当温度上升到 左右时,溶液中的 种脱氧核苷酸在耐高温的 DNA 聚合酶的作用下,根据碱基互补配对原则合成新的 DNA 链。
第一轮循环的产物作为第二轮反应的模板,经过变性、复性和延伸三步产生第二轮循环的产物。第二轮循环的产物作为第三轮反应的模板,经过变性、复性和延伸三步产生第三轮循环的产物。上述过程可以在 PCR 扩增仪(PCR 仪)中自动完成,完成以后,常采用琼脂糖凝胶电泳来鉴定 PCR 的产物。
真核细胞和细菌的 DNA 聚合酶都需要 激活。因此,PCR 反应缓冲液中一般要添加 。引物是一小段能与 DNA 母链的一段碱基序列互补配对的短单链核酸。用于 PCR 的引物长度通常为 个核苷酸。
迄今为止,在几乎所有生物体内发现的 DNA 聚合酶都不能以单个脱氧核苷酸为起点,将游离的脱氧核苷三磷酸从头聚合形成多核苷酸链,而只能将脱氧核苷酸掺入到一段已存在的核苷酸链(即引物)上。就大多数生物而言,引物通常为6~8个碱基长度的单链 RNA 分子。
由于引物是根据碱基互补配对原理(即形成氢键)"搭在"DNA 模板特定区域上的(该过程称为"退火"),因此,在较低的温度下,引物可能会与不完全配对的 DNA 模板区域结合,造成 PCR 扩增产物的多样化(即非特异性)。此外,随着扩增轮数的递增,DNA 扩增产物的浓度(也包括黏度)急剧上升,严重影响 PCR系统中的分子扩散,直至过早终止扩增反应。因此,PCR 的连续多轮循环必须在高温条件下进行,而 PCR 技术的核心便是耐热 DNA 聚合酶的发现,此类耐热酶大都来自生活在温泉中的嗜热细菌。
一般而言,衡量 PCR 的质量指标包含:(1)特异性:扩增产物纯净单一,引物长度和退火温度是关键影响因素。(2)准确性:扩增产物的序列忠实于模板 DNA序列,与所用高温聚合酶的保真性有关。(3)有效性:扩增产物高产量,与酶量和镁离子浓度有关。
三、切割和连接是构建表达载体的主要方式
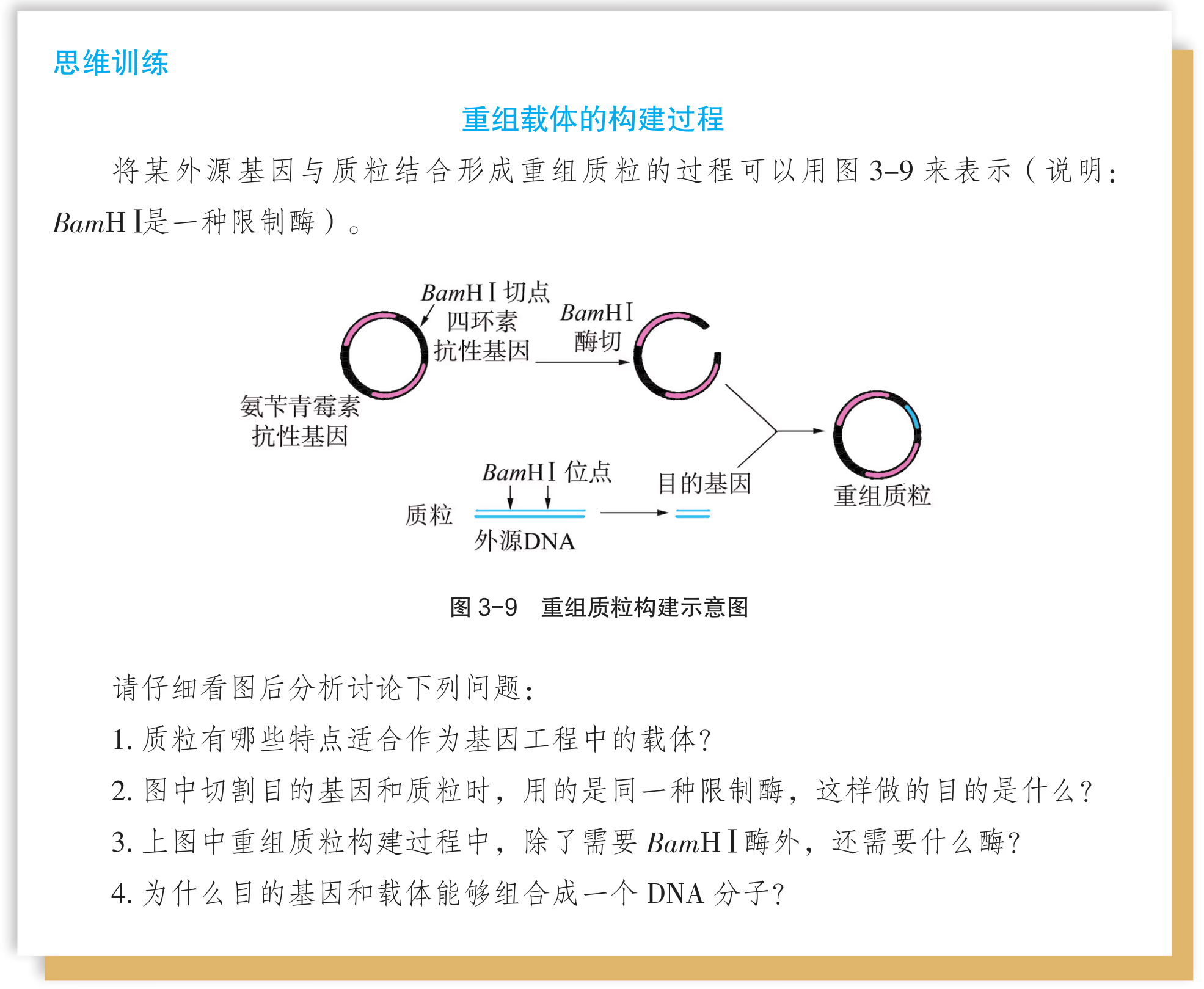
将 PCR 扩增获得的目的基因和合适的载体用相应的限制性内切核酸酶切割出黏性末端(简称"切"),然后加入 DNA 连接酶将两者连接为一体(简称"接"),形成含目的基因且能表达的表达载体,该过程称为表达载体的构建。
如果目的基因和载体均用相同的限制性内切核酸酶切开,那么它们都带有相同的黏性末端,因而可在较低温度下(一般在 4~26 ℃)"粘"在一起,并在 DNA 连接酶的作用下封闭所有的缺口,成为稳定的重组 DNA 分子。但在此过程中,需要注意下列问题:
- 由于目的基因两侧的黏性末端相同,因而能以正反两种方式等概率接入载体,但在基因工程大多数应用中,只有一种方式是可用的。
- 由于载体片段两侧的黏性末端也相同,因而在连接时载体两个黏性末端可以自我环化,导致载体"空载"(即非重组 DNA分子)。
- 根据 DNA 连接酶的工作原理,目的基因和载体 DNA 只需黏性末端相同便能有效连接,也就是说,两种 DNA并不必须用同一种酶切开。
- 如果使用两种识别序列不同、但黏性末端相同的限制性内切核酸酶分别切开目的基因和载体 DNA,那么两者连接后原来的限制性内切核酸酶识别序列会消失,也就是说,不能用相应的限制性内切核酸酶从重组 DNA 分子中重新"卸下"目的基因。
获取了足够量的 Bt 基因后,下一步就是要让基因在受体细胞中稳定存在,并且遗传给下一代;同时,使 Bt 基因能够表达和发挥作用,这就需要构建基因表达载体。这一步是培育转基因抗虫棉的核心工作。
基因表达载体是载体的一种,除目的基因、标记基因外,它还必须有启动子(promoter)、终止子(terminator)等。启动子是一段有特殊序列结构的 DNA 片段,位于基因的上游,紧挨转录的起始位点,它是 RNA 聚合酶识别和结合的部位,有了它才能驱动基因转录出 mRNA,最终表达出人类需要的蛋白质。有时为了满足应用需要,会在载体中人工构建诱导型启动子,当诱导物存在时,可以激活或抑制目的基因的表达。终止子相当于一盏红色信号灯,使转录在所需要的地方停下来,它位于基因的下游,也是一段有特殊序列结构的 DNA 片段。
在构建抗虫棉的基因表达载体时,首先会用一定的限制酶切割载体,使它出现一个切口;然后用同种限制酶或能产生相同末端的限制酶切割含有目的基因的 DNA 片段;再利用 DNA 连接酶将目的基因片段拼接到载体的切口处,这样就形成了一个重组 DNA 分子(图 3-6)。
四、将 DNA 分子导入受体细胞
经连接操作后的 DNA 分子只有导入受体细胞,才能借助细胞内的基因表达系统使目的基因所蕴含的遗传信息得以表达。受体细胞可以是微生物、植物或动物细胞,完全由基因工程的应用性质而定。例如,采用基因工程技术改良农作物的性状,受体细胞显然来自相应的植物。
在自然条件下,DNA 很难进入细胞。为了大幅度提高 DNA 分子进入受体细胞的效率,通常需要对受体细胞进行特殊的处理,这一操作称为转化(简称"转")。转化的方法可以采用物理方法(如电击法)、化学方法(如氯化钙法)、生物方法(如农杆菌法)等。即使如此,转化成功的效率也只有 ,因而需要高效筛选出接纳了 DNA 分子的受体细胞。为了做到这一点,转化后的细胞需要经过一段时间的培养,使得用于筛选的性状得以表达,这个过程称为扩增(简称"增")。

构建好的基因表达载体需要通过一定的方式才能进入受体细胞。我国科学家采用他们独创的一种方法------花粉管通道法,将 Bt 基因导入了棉花细胞。花粉管通道法有多种操作方式。例如,可以用微量注射器将含目的基因的 DNA 溶液直接注入子房中;可以在植物受粉后的一定时间内,剪去柱头,将 DNA 溶液滴加在花柱切面上,使目的基因借助花粉管通道进入胚囊。除此之外,将目的基因导入植物细胞常用的方法还有农杆菌转化法。
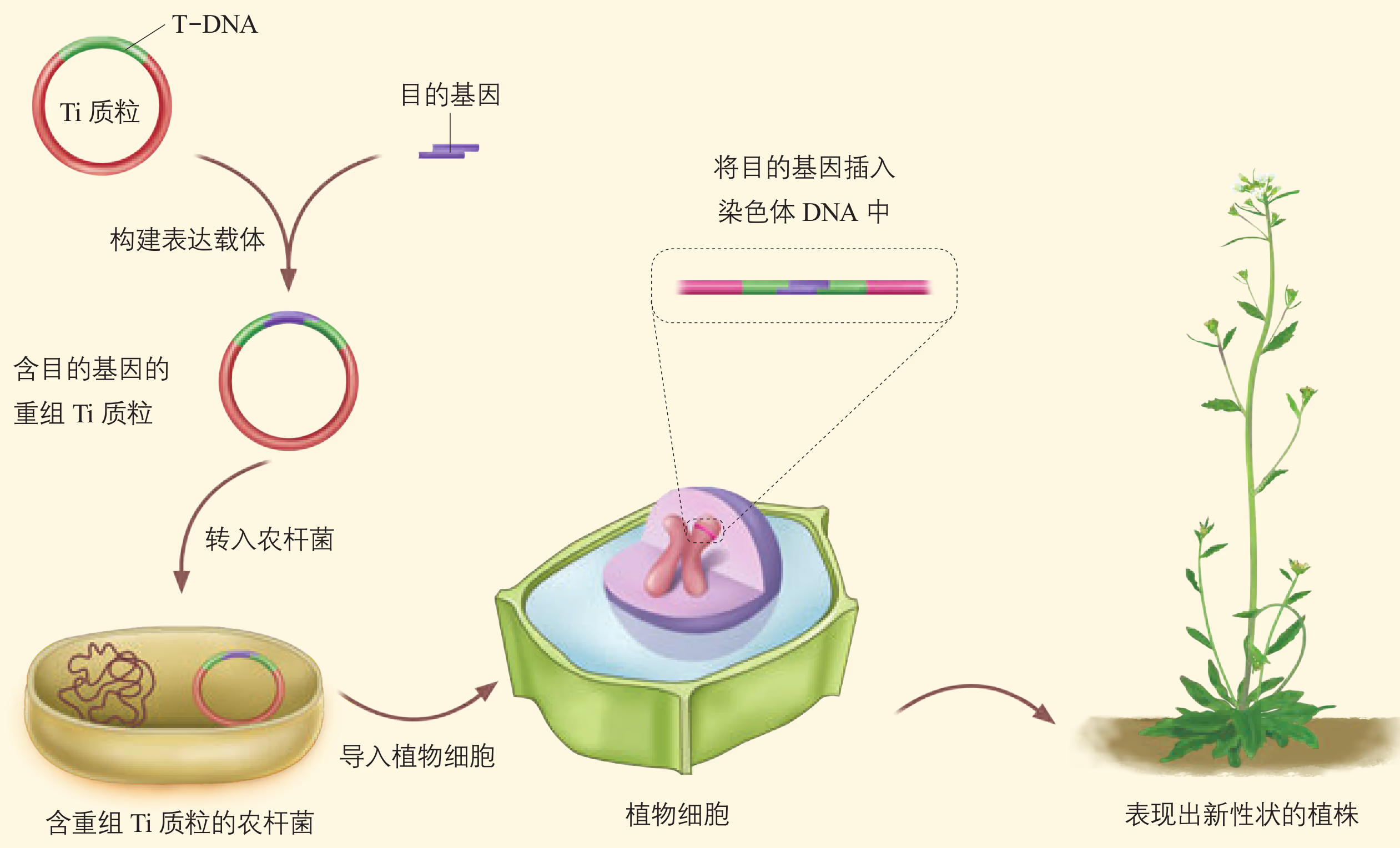
农杆菌转化法:转化(transformation)是指目的基因进入受体细胞内,并且在受体细胞内维持稳定和表达的过程。
农杆菌是一种在土壤中生活的微生物,能在自然条件下侵染双子叶植物和裸子植物,而对大多数单子叶植物没有侵染能力。农杆菌细胞内含有 Ti 质粒,当它侵染植物细胞后,能将 Ti 质粒上的 T-DNA(可转移的 DNA)转移到被侵染的细胞,并且将其整合到该细胞的染色体 DNA 上。根据农杆菌的这种特点,如果将目的基因插入 Ti 质粒的 T-DNA 中,通过农杆菌的转化作用,就可以使目的基因进入植物细胞。
根据受体植物的不同,所用的具体转化方法有所区别。例如,可以将新鲜的从叶片上取下的圆形小片与农杆菌共培养,然后筛选转化细胞,并再生成植株;可以将花序直接浸没在含有农杆菌的溶液中一段时间,然后培养植株并获得种子,再进行筛选、鉴定等。随着转化方法的突破,用农杆菌侵染水稻、玉米等多种单子叶植物也取得了成功。
五、借助标记基因筛选和鉴定含目的基因的受体细胞
由于在转化操作中,只有少数细胞能接纳 DNA 分子,而且进入受体细胞的 DNA 分子还有可能含有空载质粒。因此,需要借助有效的实验技术快速、准确地找到正确导入(甚至高效表达)目的基因的受体细胞,这一过程称为筛选与鉴定(简称"检")。
筛选含目的基因的受体细胞常用抗药性筛选法和显色筛选法。抗药性筛选法实施的前提条件是载体 DNA 携带抗生素的抗性基因。例如,大肠杆菌 pBR322 质粒含有氨苄青霉素抗性基因()和四环素抗性基因(),这些可供筛选使用的功能性基因称为标记基因。如果外源 DNA 插在质粒 pBR322 的 BamH Ⅰ 位点处,则只需将转化扩增的细菌涂布在含有氨苄青霉素(Amp)的固体平板上,理论上能长出的菌落便是转化成功的克隆(图 3-13)。上述筛选获得的转化成功克隆中有两种类型,即含重组质粒的克隆和含空载质粒的克隆。为了进一步区分这两类克隆,需要采用图 3-13 中(B)所示的方法进行第二轮筛选。由于外源 DNA 片段在 BamH Ⅰ 位点的重组导致载体 DNA 的 基因插入灭活,含重组质粒的克隆呈 表型(上标"r"代表抗性;"s"代表敏感);而含空载质粒的克隆则为 表型。因此,含重组质粒的克隆只能在 Amp 平板上生长;而含空载质粒的克隆可出现在同时含 Amp 和 Tet 的平板上。
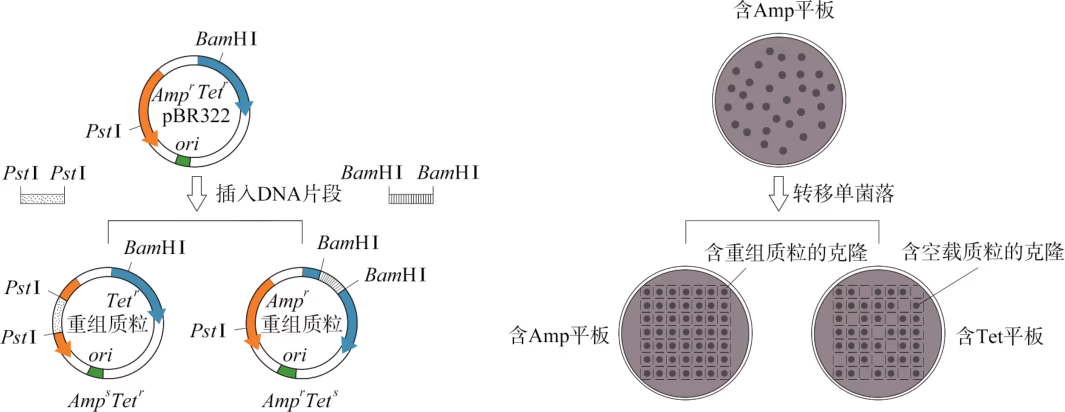
很多大肠杆菌的质粒上含有 lacZ' 标记基因,其表达的酶蛋白可将一种无色的化合物(X-gal)水解成蓝色产物。重组 DNA 技术常用的大肠杆菌质粒 pUC18/19 同时携带 和 lacZ' 两个标记基因。将外源 DNA 插在 lacZ' 标记基因内部的任何限制性内切核酸酶切割位点处,将转化扩增的细菌涂布在同时含有 Amp 和 X-gal 的固体培养基上,在长出来的克隆中,白色克隆即为含重组质粒的克隆(图 3-14)。
除了上述两大类标记基因外,在产业化生产用于食品和药品的基因工程产品时,出于安全考虑,还需要使用其他无害的标记基因以及相应的筛选方法。经抗药性或显色筛选获得的含目的基因的受体细胞还需进一步鉴定重组在载体上的目的基因位置是否正确以及是否能高效表达,这需要使用 PCR 技术和基因表达产物(蛋白质)的结构和功能分析技术。
六、构建转基因动植物需要针对特定受体细胞进行操作
如果构建多细胞的转基因动植物,还需将表达载体导入可发育成动植物个体的受体细胞中,例如动物的生殖细胞(精细胞和卵细胞)、受精卵(或早期胚胎细胞)、诱导性多能干细胞等,植物的愈伤组织(或其他组织的原生质体)、生殖细胞或胚胎组织等。DNA 显微注射是动物细胞基因转移普遍采用的一种物理转化方法,其基本操作程序如下:通过激素疗法使雌鼠超数排卵,并与雄性小鼠交配后从其输卵管内取出受精卵,用吸管将受精卵固定在倒置显微镜上,然后借助玻璃注射针向受精卵注入 DNA 溶液;将注射了 DNA 的受精卵移植到母鼠子宫中发育,继而繁殖转基因小鼠子代。此外,当采用动物病毒(如腺相关病毒和慢病毒)DNA 作表达载体时,可以通过病毒感染的方式将目的基因高效导入动物细胞中。
植物细胞的常用转化方法是用含有表达载体的根瘤农杆菌感染受体细胞。此外,也可用含有表达载体的植物病毒直接感染植物的叶或茎等组织,这些病毒能将目的基因迅速扩散至整株植物。由于植物的愈伤组织能再生出整株植物,因此转基因植物的构建要比转基因动物的构建更加简便。转基因动植物培育出来后,仍需要使用 PCR 等技术确认转基因是否获得成功,以及目的基因在动植物体内是否正确表达。
目的基因进入受体细胞后,是否稳定维持和表达其遗传特性,只有通过检测与鉴定才能知道,这也是检查转基因抗虫棉是否培育成功的一步。首先是分子水平的检测,包括通过 PCR 等技术检测棉花的染色体 DNA 上是否插入了 Bt 基因或检测 Bt 基因是否转录出了 mRNA;从转基因棉花中提取蛋白质,用相应的抗体进行抗原---抗体杂交,检测 Bt 基因是否翻译成 Bt 抗虫蛋白等。其次,还需要进行个体生物学水平的鉴定。例如,通过采摘抗虫棉的叶片饲喂棉铃虫来确定 Bt 基因是否赋予了棉花抗虫特性以及抗性的程度。
在获得转基因产品的过程中,还可以通过构建基因文库来获取目的基因,并且由于不同转基因产品所需要的目的基因不同,受体细胞又有植物、动物、微生物之分,因此基因表达载体的构建方法不是千篇一律的,将目的基因导入受体细胞的方法也不是完全相同的。例如,将目的基因导入动物受精卵最常用的一种方法是利用显微注射将目的基因注入动物的受精卵中(图 3-8),这个受精卵将发育成为具有新性状的动物。
在基因工程操作中,常用原核生物作为受体细胞,其中以大肠杆菌应用最为广泛。研究人员一般先用 处理大肠杆菌细胞,使细胞处于一种能吸收周围环境中 DNA 分子的生理状态,然后再将重组的基因表达载体导入其中。目前,基因工程已经步入产业化发展阶段,具有巨大的生产力和发展潜力,我们在充满信心和期待的同时,还要严格规范操作流程,科学、客观地评估风险,只有这样,才能让基因工程永葆生机。